

The brain on stress: vulnerability and plasticity of the prefrontal cortex over the life course
- PMID: 23849196
- PMCID: PMC3753223
- DOI: 10.1016/j.neuron.2013.06.028
The brain on stress: vulnerability and plasticity of the prefrontal cortex over the life course
Abstract
The prefrontal cortex (PFC) is involved in working memory and self-regulatory and goal-directed behaviors and displays remarkable structural and functional plasticity over the life course. Neural circuitry, molecular profiles, and neurochemistry can be changed by experiences, which influence behavior as well as neuroendocrine and autonomic function. Such effects have a particular impact during infancy and in adolescence. Behavioral stress affects both the structure and function of PFC, though such effects are not necessarily permanent, as young animals show remarkable neuronal resilience if the stress is discontinued. During aging, neurons within the PFC become less resilient to stress. There are also sex differences in the PFC response to stressors. While such stress and sex hormone-related alterations occur in regions mediating the highest levels of cognitive function and self-regulatory control, the fact that they are not necessarily permanent has implications for future behavior-based therapies that harness neural plasticity for recovery.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
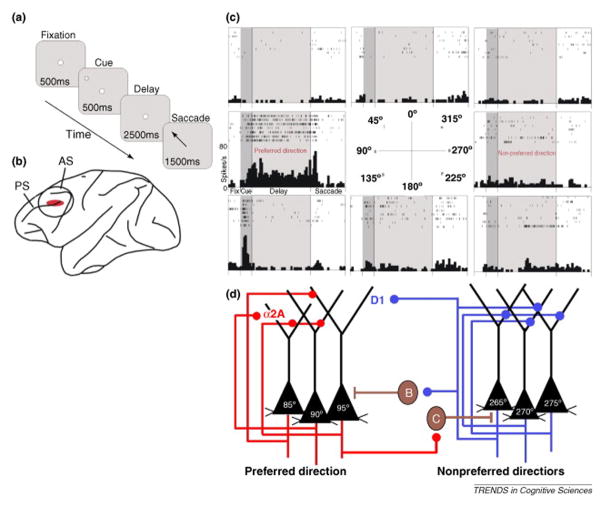
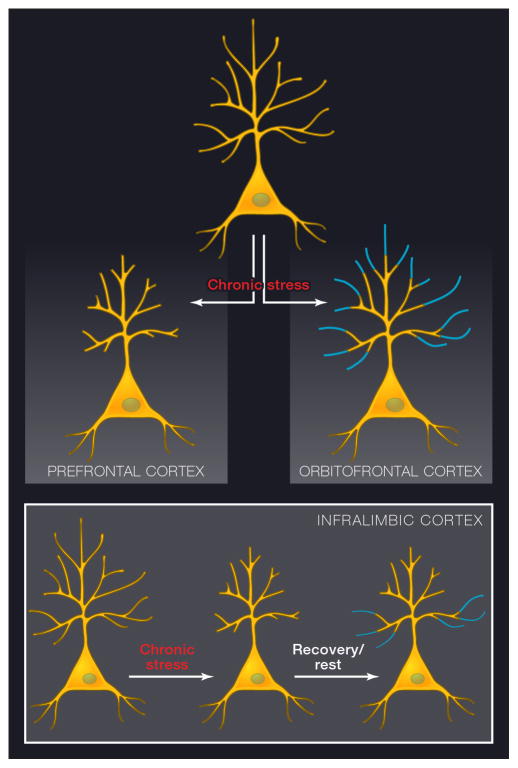
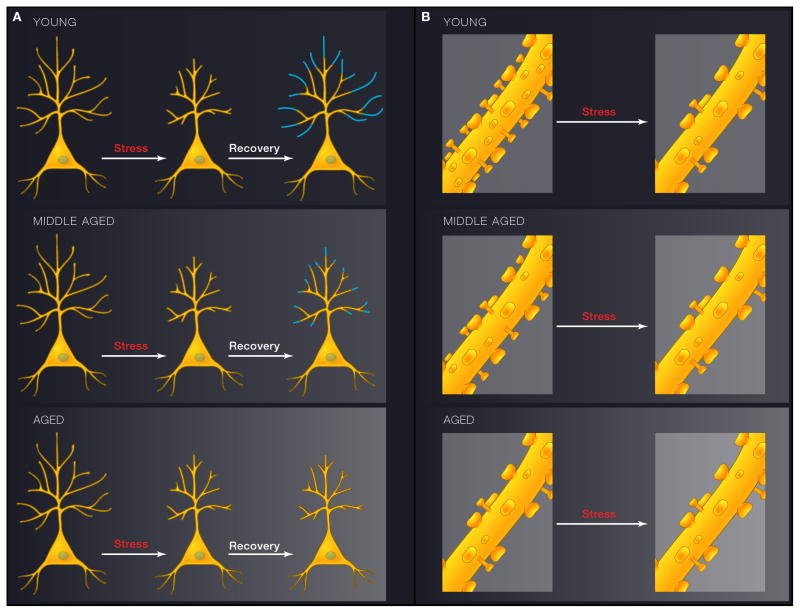
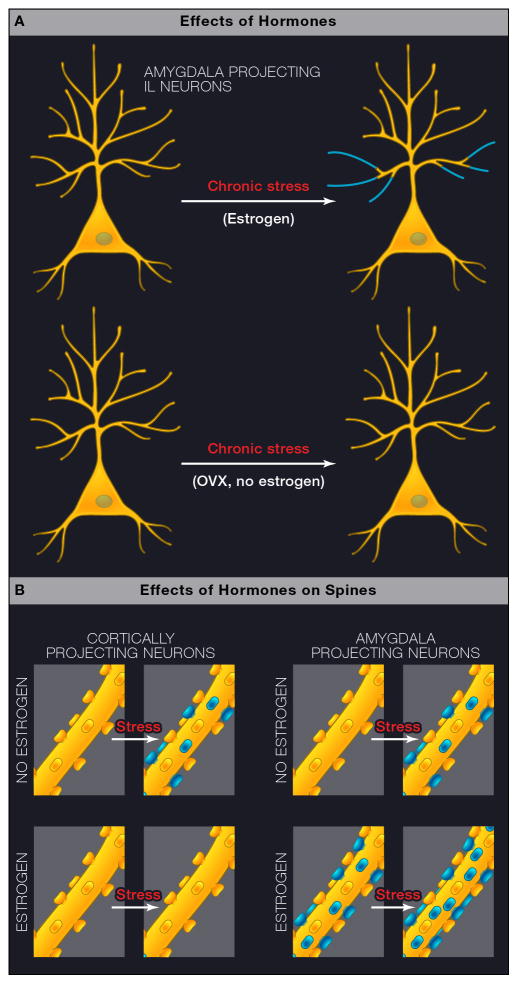
Comment in
-
An Overly Permissive Extension.Perspect Psychol Sci. 2016 Jul;11(4):442-50. doi: 10.1177/1745691616635593. Perspect Psychol Sci. 2016. PMID: 27474132
References
-
- Ahima R, Krozowski Z, Harlan R. Type I corticosteroid receptor-like immunoreactivity in the rat CNS: distribution and regulation by corticosteroids. J Comp Neurol. 1991;313:522–538. - PubMed
-
- Ahima RS, Harlan RE. Charting of type II glucocorticoid receptor-like immunoreactivity in the rat central nervous system. Neuroscience. 1990;39:579–604. - PubMed
-
- Anda RF, Butchart A, Felitti VJ, Brown DW. Building a framework for global surveillance of the public health implications of adverse childhood experiences. Am J Prev Med. 2010;39:93–98. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
