

Division of labor between the chromodomains of HP1 and Suv39 methylase enables coordination of heterochromatin spread
- PMID: 23849629
- PMCID: PMC3752401
- DOI: 10.1016/j.molcel.2013.06.013
Division of labor between the chromodomains of HP1 and Suv39 methylase enables coordination of heterochromatin spread
Abstract
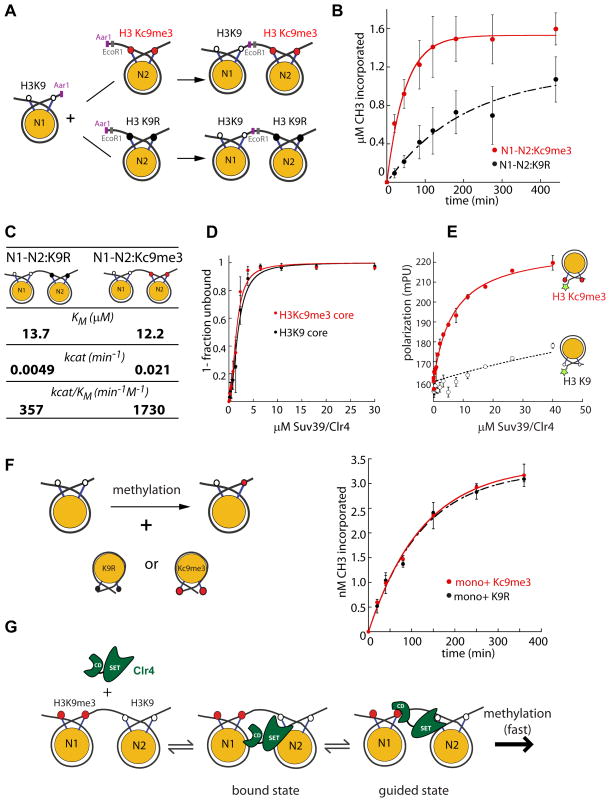
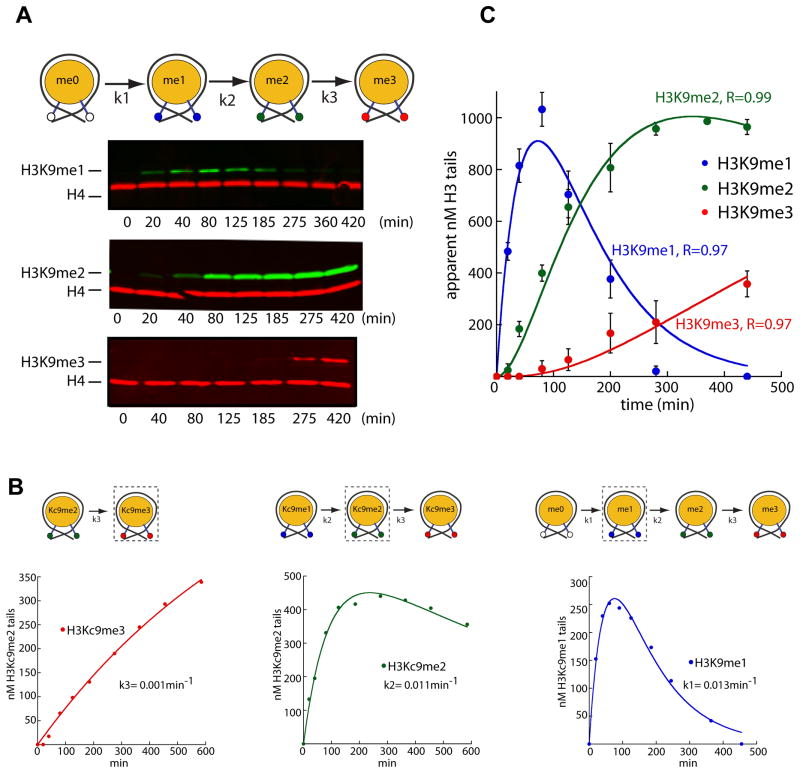
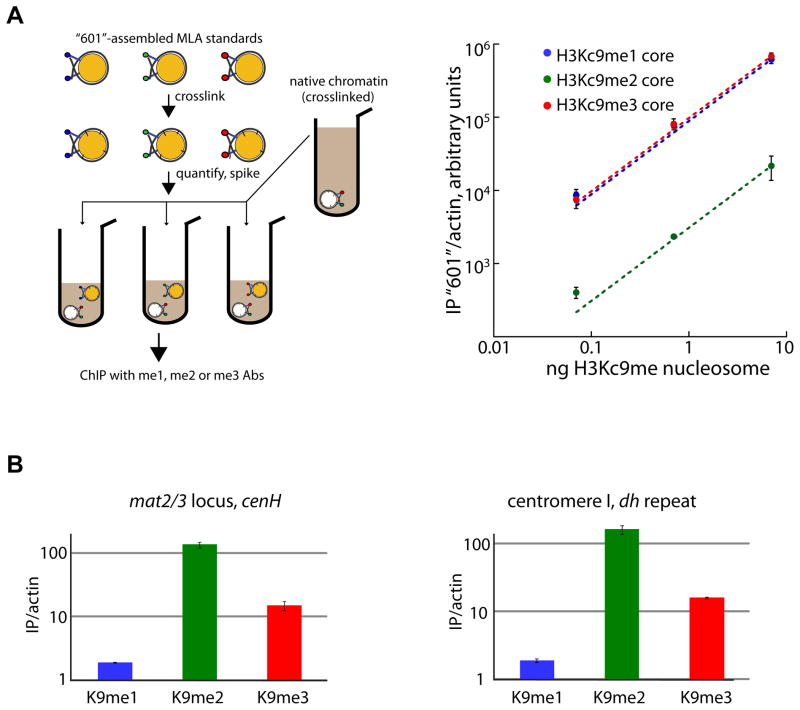
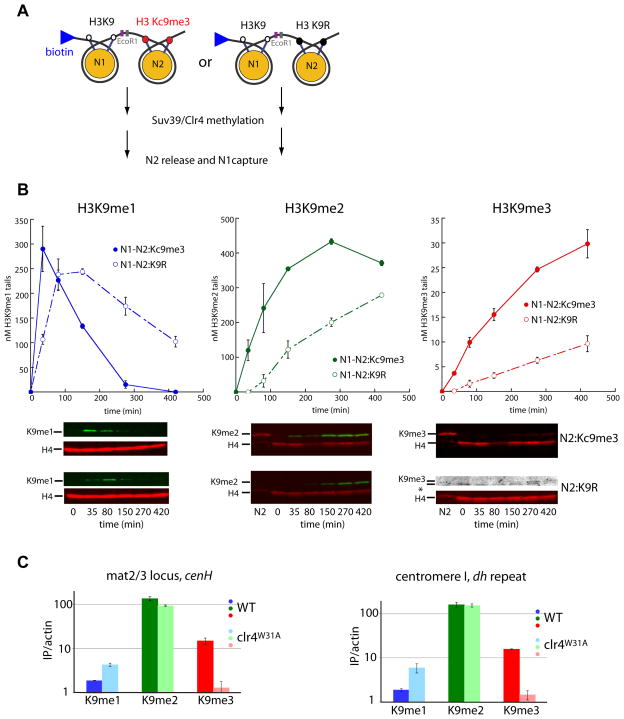
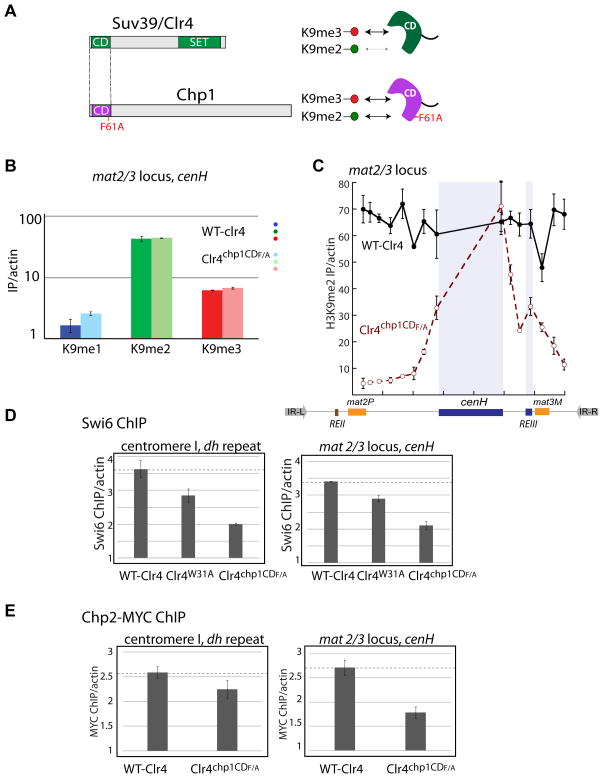
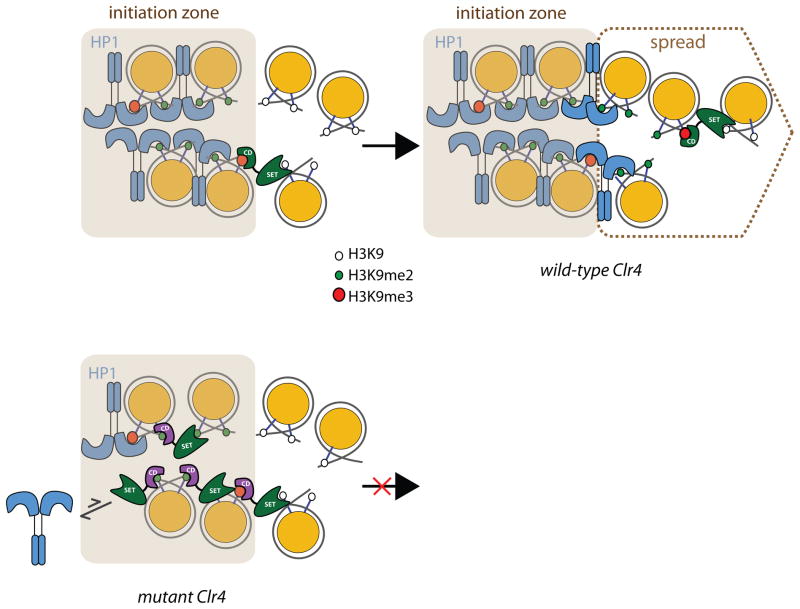
In Schizosaccharomyces pombe, heterochromatin spread, which is marked by histone 3 lysine 9 methylation (H3K9me), requires the chromodomains (CDs) of the H3K9 methylase Suv39/Clr4 and the HP1/Swi6 protein. It is unclear how the actions of these two H3K9me-recognizing CDs are coordinated. We find that the intrinsic preference of Suv39/Clr4 is to generate dimethylated H3K9 product. The recognition of pre-existing H3K9me marks by the CD of Suv39/Clr4 stimulates overall catalysis, enabling the accumulation of small amounts of trimethylated product in vivo. Coincidentally, the Suv39/Clr4 CD, unlike the HP1/Swi6 CD, has been shown to prefer the trimethyl state over the dimethyl state. We show that this preference enables efficient heterochromatin spread in vivo by reducing competition with HP1 proteins for the more prevalent dimethyl state. Our results reveal a strategy by which "writers" and "readers" of a chromatin mark exploit different methylation states on the same residue in order to facilitate collaboration and avoid competition.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Bhalla KN. Epigenetic and Chromatin Modifiers As Targeted Therapy of Hematologic Malignancies. J Clin Oncol. 2005;23:3971–3993. - PubMed
-
- Buhler M, Haas W, Gygi SP, Moazed D. RNAi-dependent and -independent RNA turnover mechanisms contribute to heterochromatic gene silencing. Cell. 2007;129:707–721. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
