

A virocentric perspective on the evolution of life
- PMID: 23850169
- PMCID: PMC4326007
- DOI: 10.1016/j.coviro.2013.06.008
A virocentric perspective on the evolution of life
Abstract
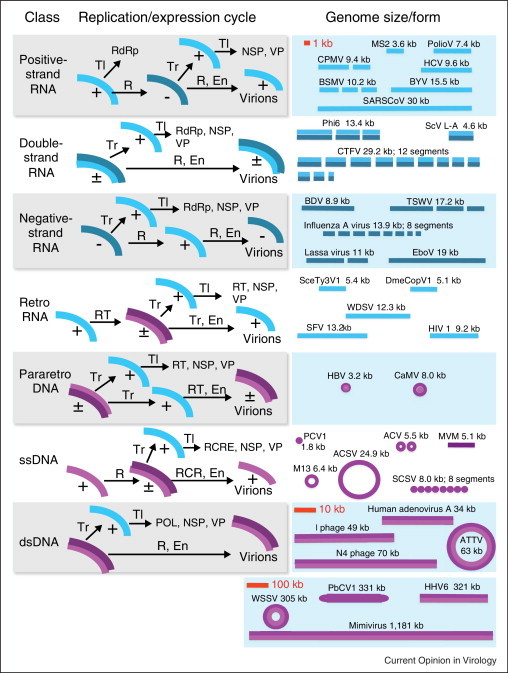
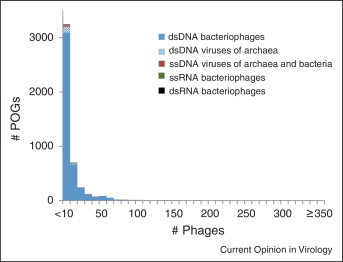
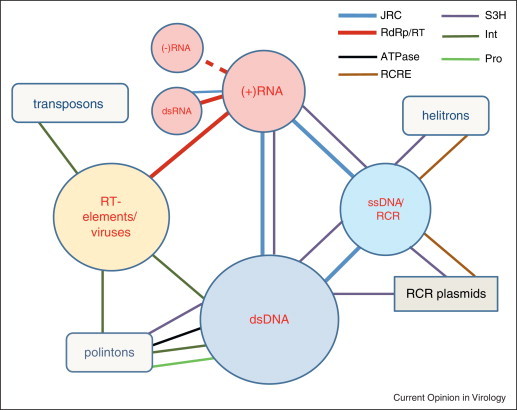
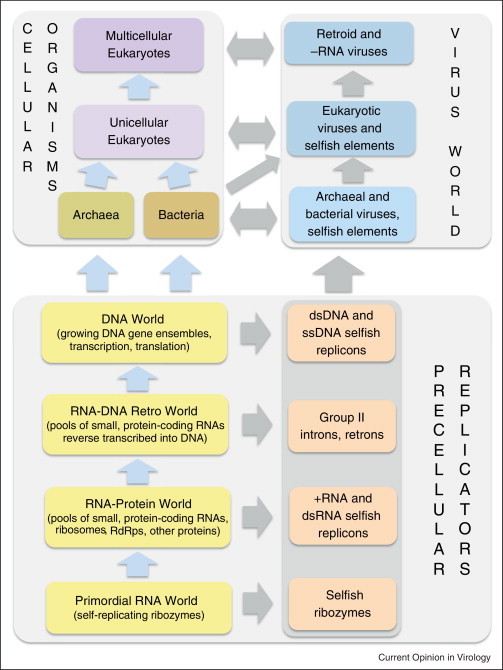
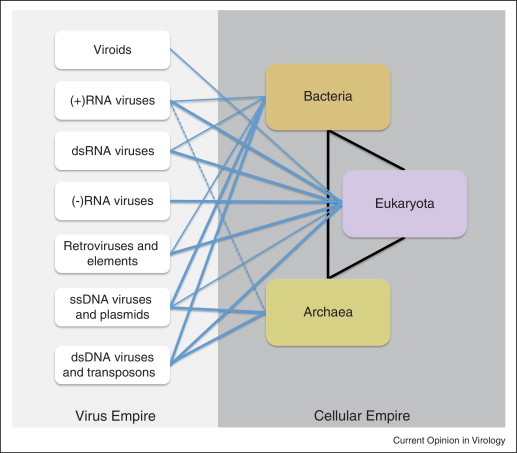
Viruses and/or virus-like selfish elements are associated with all cellular life forms and are the most abundant biological entities on Earth, with the number of virus particles in many environments exceeding the number of cells by one to two orders of magnitude. The genetic diversity of viruses is commensurately enormous and might substantially exceed the diversity of cellular organisms. Unlike cellular organisms with their uniform replication-expression scheme, viruses possess either RNA or DNA genomes and exploit all conceivable replication-expression strategies. Although viruses extensively exchange genes with their hosts, there exists a set of viral hallmark genes that are shared by extremely diverse groups of viruses to the exclusion of cellular life forms. Coevolution of viruses and host defense systems is a key aspect in the evolution of both viruses and cells, and viral genes are often recruited for cellular functions. Together with the fundamental inevitability of the emergence of genomic parasites in any evolving replicator system, these multiple lines of evidence reveal the central role of viruses in the entire evolution of life.
Published by Elsevier B.V.
Figures
References
-
- Bergh O., Borsheim K.Y., Bratbak G., Heldal M. High abundance of viruses found in aquatic environments. Nature. 1989;340:467–468. - PubMed
-
- Rosario K., Breitbart M. Exploring the viral world through metagenomics. Curr Opin Virol. 2011;1:289–297. - PubMed
-
An overview of the discovery of new viruses and virus–host associations through analysis of metagenomic data.
-
- Culley A.I. Virophages to viromes: a report from the frontier of viral oceanography. Curr Opin Virol. 2011;1:52–57. - PubMed
-
An overview of the discovery of novel marine viruses, including giant viruses and virophages, and their roles in the ecology of the oceans.
-
- Rohwer F. Global phage diversity. Cell. 2003;113:141. - PubMed
-
- Colson P., de Lamballerie X., Fournous G., Raoult D. Reclassification of giant viruses composing a fourth domain of life in the new order Megavirales. Intervirology. 2012;55:321–332. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
