

Motor cortex feedback influences sensory processing by modulating network state
- PMID: 23850595
- PMCID: PMC3742632
- DOI: 10.1016/j.neuron.2013.06.008
Motor cortex feedback influences sensory processing by modulating network state
Abstract
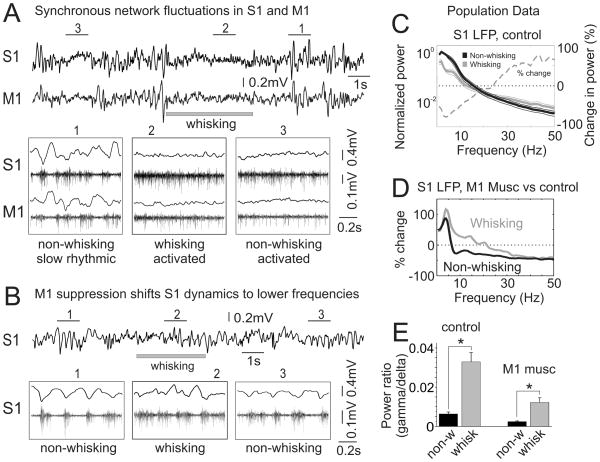
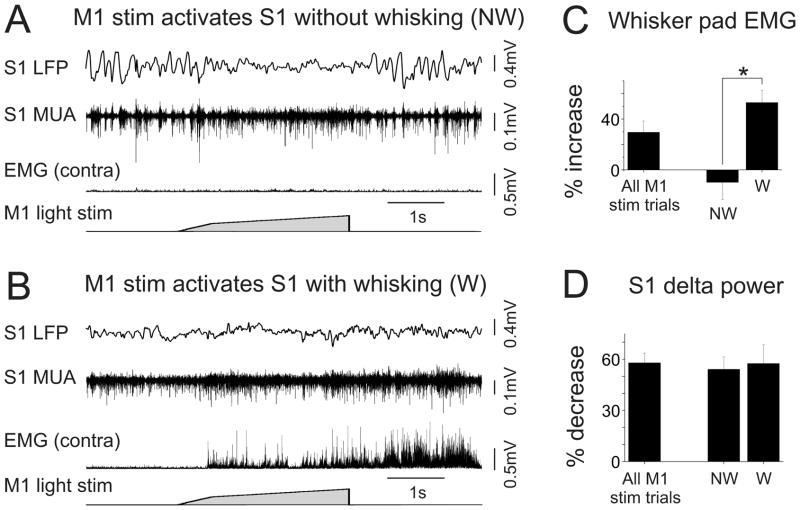
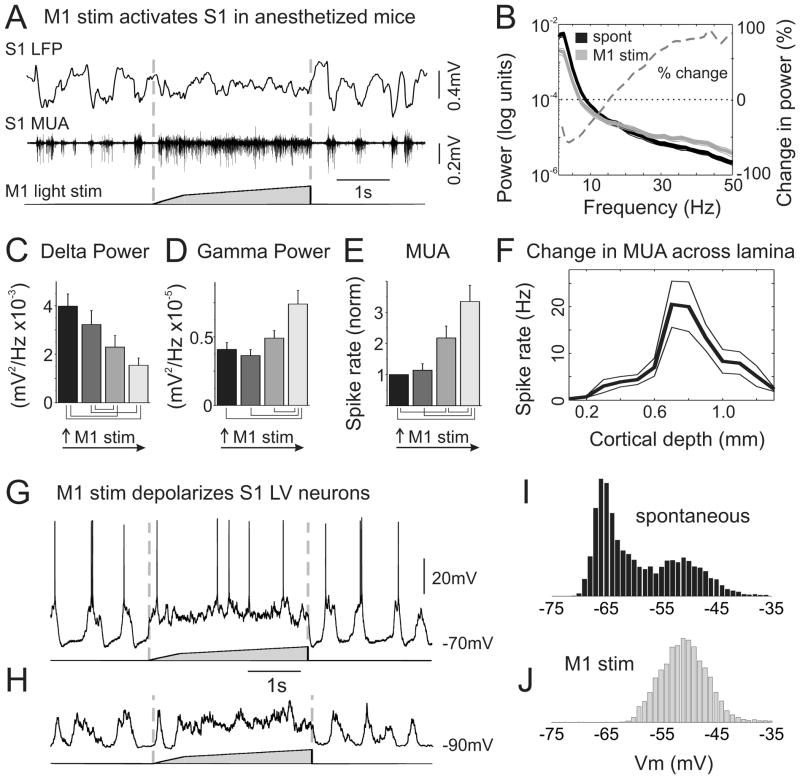
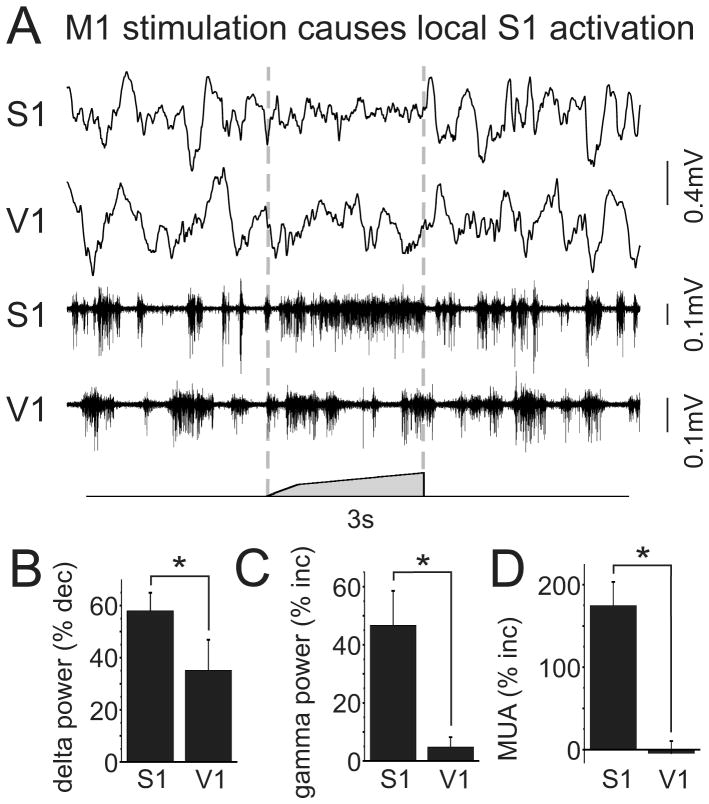
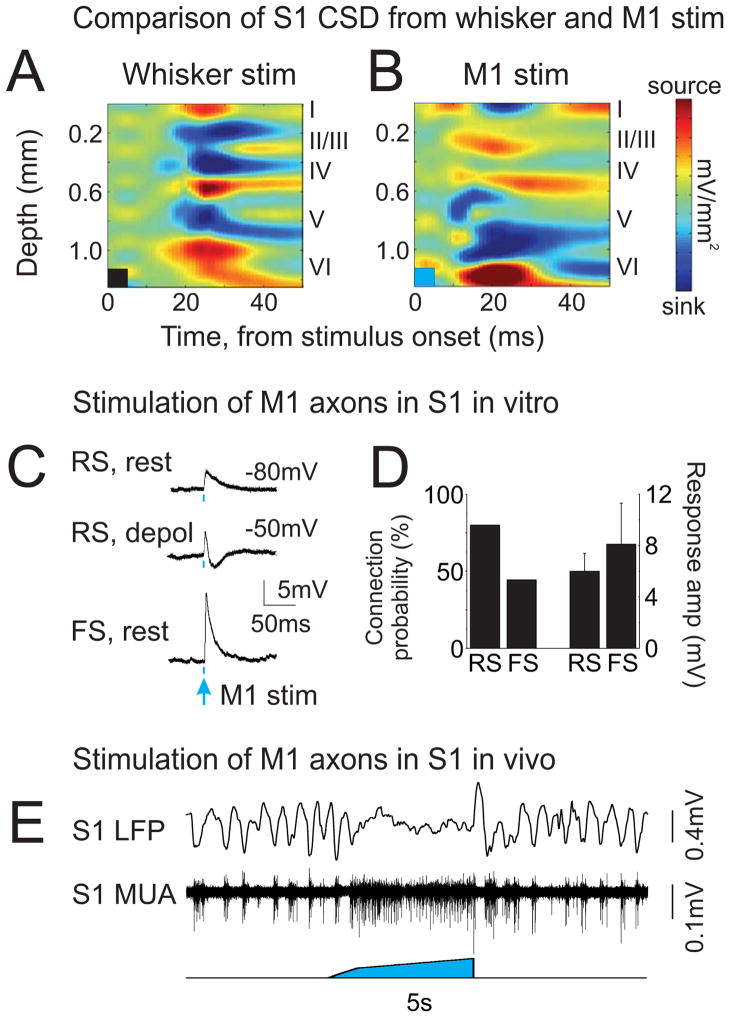
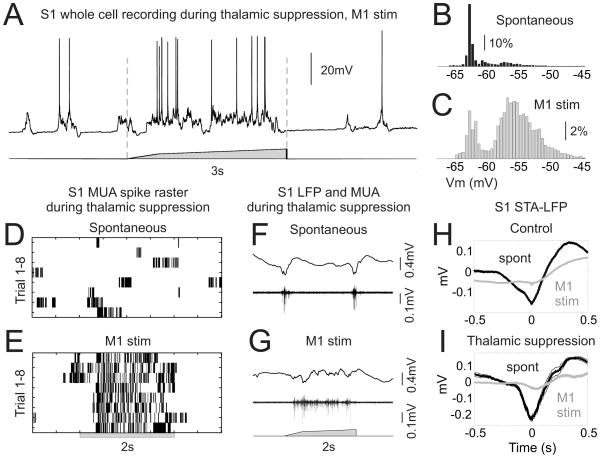
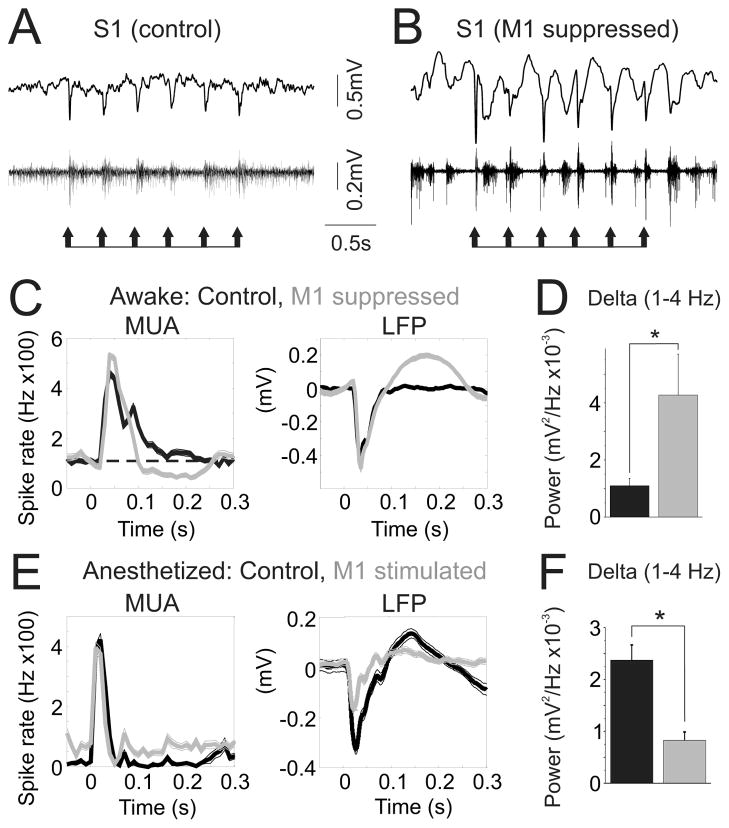
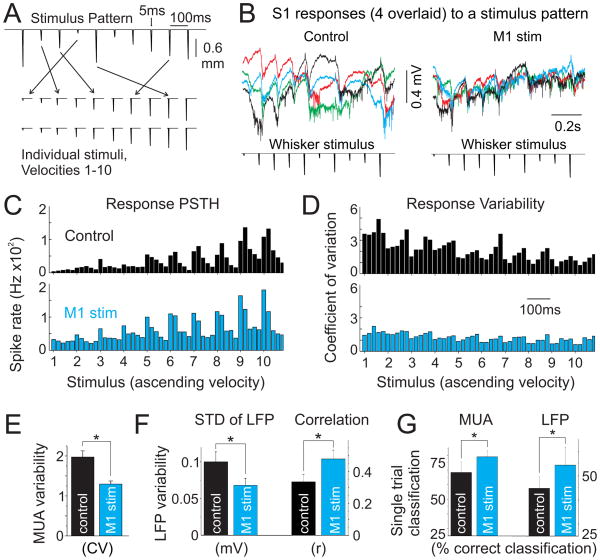
Long-range corticocortical communication may have important roles in context-dependent sensory processing, yet we know very little about how these pathways influence their target regions. We studied the influence of primary motor cortex activity on primary somatosensory cortex in the mouse whisker system. We show that primary motor and somatosensory cortices undergo coherent, context-dependent changes in network state. Moreover, we show that motor cortex activity can drive changes in somatosensory cortex network state. A series of experiments demonstrate the involvement of the direct corticocortical feedback pathway, providing temporally precise and spatially targeted modulation of network dynamics. Cortically mediated changes in network state significantly impact sensory coding, with activated states increasing the reliability of responses to complex stimuli. By influencing network state, corticocortical communication from motor cortex may ensure that during active exploration the relevant sensory region is primed for enhanced sensory discrimination.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Top-down control of cortical state.Neuron. 2013 Aug 7;79(3):408-10. doi: 10.1016/j.neuron.2013.07.034. Neuron. 2013. PMID: 23931991 Free PMC article.
References
-
- Arieli A, Sterkin A, Grinvald A, Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–1871. - PubMed
-
- Berger H. Ueber das Elektroenkephalogramm des Menschen. Arch Psychiatr Nervenkrankh. 1929;87:527–570.
-
- Brecht M. Movement, confusion, and orienting in frontal cortices. Neuron. 2011;72:193–196. - PubMed
-
- Buzsaki G. Rhythms of the brain. New York: Oxford University Press; 2006.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
