

Hematopoietic-specific Rho GTPases Rac2 and RhoH and human blood disorders
- PMID: 23850828
- PMCID: PMC3997055
- DOI: 10.1016/j.yexcr.2013.07.002
Hematopoietic-specific Rho GTPases Rac2 and RhoH and human blood disorders
Abstract
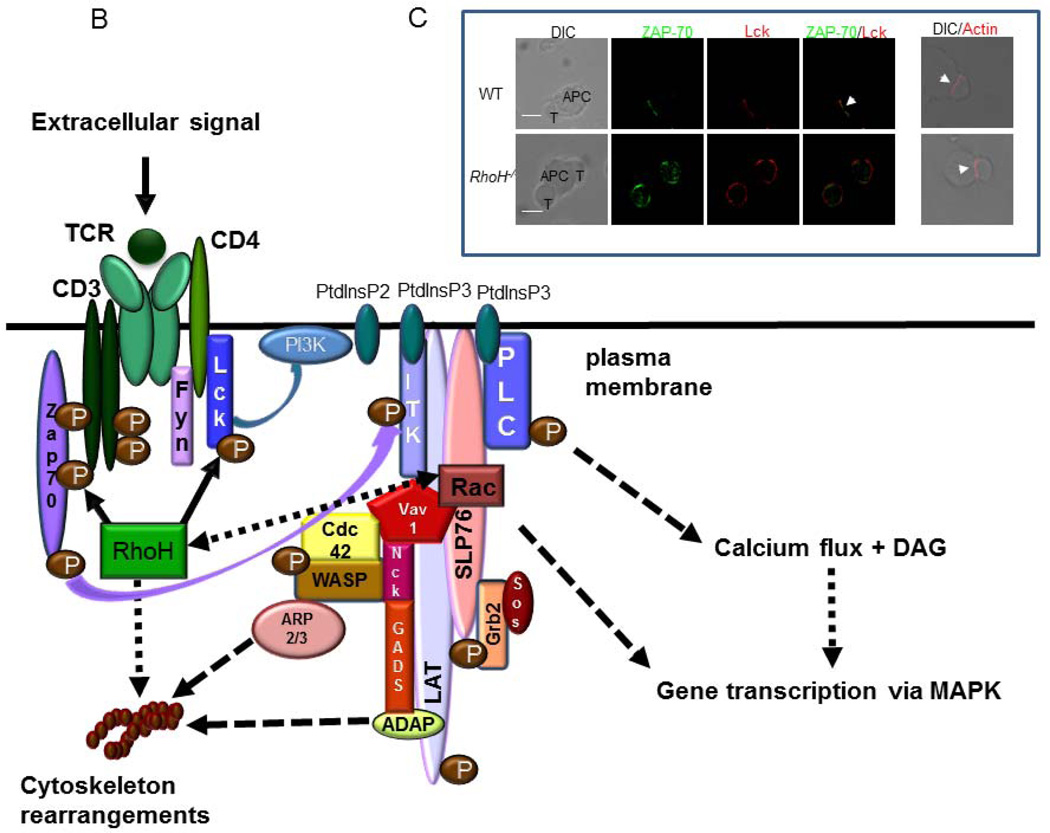
The small guanosine triphosphotases (GTPases) Rho proteins are members of the Ras-like superfamily. Similar to Ras, most Rho GTPases cycle between active GTP-bound, and inactive GDP-bound conformations and act as molecular switches that control multiple cellular functions. While most Rho GTPases are expressed widely, the expression of Rac2 and RhoH are restricted to hematopoietic cells. RhoH is an atypical GTPase that lacks GTPase activity and remains in the active conformation. The generation of mouse knock-out lines has led to new understanding of the functions of both of these proteins in blood cells. The phenotype of these mice also led to the identification of mutations in human RAC2 and RHOH genes and the role of these proteins in immunodeficiency diseases. This review outlines the basic biology of Rho GTPases, focusing on Rac and RhoH and summarizes human diseases associated with mutations of these genes.
Keywords: Cytoskeleton; Hematopoiesis; Rac; Rho GTPases; RhoH.
© 2013 Elsevier Inc. All rights reserved.
Figures


References
-
- Andersen PR, et al. Generation of BALB-MuSV and Ha-MuSC by type C virus transduction of homologous transforming genes from different species. Cell. 1981;26:129–134. - PubMed
-
- Bourne H, Sanders D, McCormick F. The GTPase superfamily: a conserved switch for diverse cell functions. Nature. 1990;348:125–132. - PubMed
-
- Aspenstrom P. The Rho GTPases have multiple effects on the actin cytoskeleton. Exp Cell Res. 1999;246:20–25. - PubMed
-
- Bourne HR, Sanders DA, McCormick F. The GTPase superfamily: conserved structure and molecular mechanism. Nature. 1991;349:117–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
