

Characterization of Neospora caninum macrophage migration inhibitory factor
- PMID: 23850997
- PMCID: PMC3805277
- DOI: 10.1016/j.exppara.2013.07.001
Characterization of Neospora caninum macrophage migration inhibitory factor
Abstract
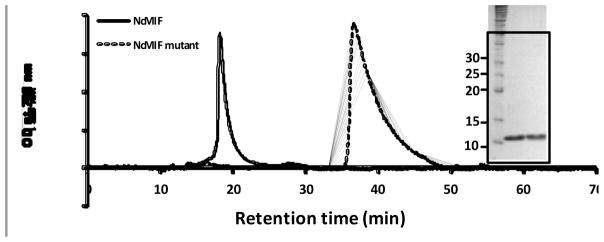
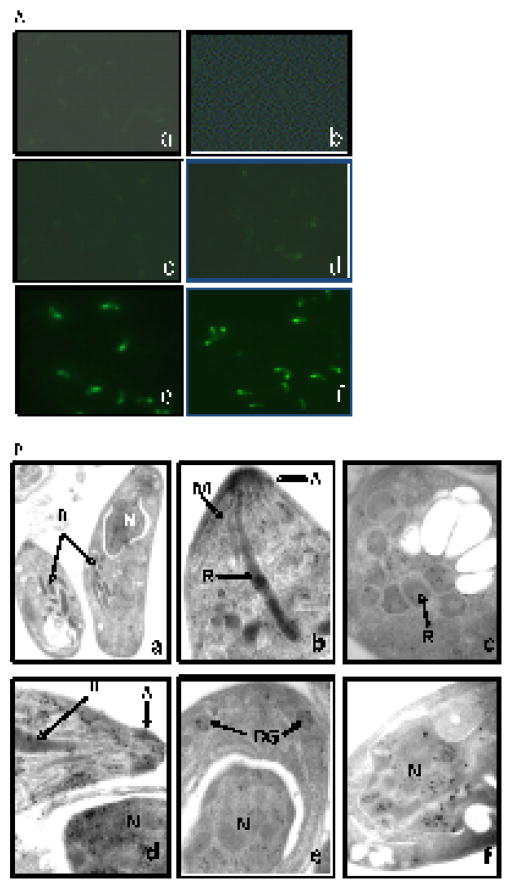
The present study is the first characterization of Neospora caninum macrophage migration inhibitory factor (NcMIF). BLAST-N analysis of NcMIF revealed high similarity (87%) to the Toxoplasma gondii MIF. NcMIF was cloned and expressed in Escherichia coli in 3 forms, NcMIF (mature protein), NcMIFm (mutation of proline-2 to glycine), and NcMIFhis (addition of a polyhistidine tag at the N-terminus). None of these recombinant NcMIFs (rNcMIF) had tautomerase, oxidoreductase, or immunologic regulatory activities. rNcMIF was unable to compete with recombinant human MIF for a MIF receptor (CD74), suggesting that NcMIF does not bind to this MIF receptor. The glycine substitution for proline-2 of NcMIF resulted in increased retention time on SEC-HPLC and decreased formation of dimers and trimers. The addition of N-terminal HIS-tag led to increased formation of trimers. Immunofluorescence staining demonstrated that NcMIF was localized to the apical end of N. caninum tachyzoites. Immunoelectron microscopy further revealed that NcMIF was present in the micronemes, rhoptries, dense granules, and nuclei. NcMIF was abundant in the tachyzoite lysate and present in excretory and secretory antigen (ESAg) preparations. Total and secretory NcMIF was more abundant in a non-pathologic clone, Ncts-8, than in the wild type isolate (NC1). Furthermore, NcMIF release by the both isolates was increased in the presence of calcium ionophore. This differential production of NcMIF by the pathologic and non-pathologic isolates of N. caninum may suggest a critical role of this molecule in the infectious pathogenesis of this parasite.
Keywords: CD74; MIF; Macrophage migration inhibitory factor; NcMIF; Neospora caninum; Tautomerase.
Published by Elsevier Inc.
Figures





References
-
- Abe R, Peng T, Sailors J, Bucala R, Metz CN. Regulation of the CTL response by macrophage migration inhibitory factor. J Immunol. 2001;166:747–753. - PubMed
-
- Aida Y, Pabst MJ. Removal of endotoxin from protein solutions by phase separation using Triton X-114. J Immunol Methods. 1990;132:191–195. - PubMed
-
- Alam A, Goyal M, Iqbal MS, Bindu S, Dey S, Pal C, Maity P, Mascarenhas NM, Ghoshal N, Bandyopadhyay U. Cysteine-3 and cysteine-4 are essential for the thioredoxin-like oxidoreductase and antioxidant activities of Plasmodium falciparum macrophage migration inhibitory factor. Free Radic Biol Med. 2011;50:1659–1668. - PubMed
-
- Arnoys EJ, Wang JL. Dual localization: proteins in extracellular and intracellular compartments. Acta Histochem. 2007;109:89–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
