

Network link prediction by global silencing of indirect correlations
- PMID: 23851447
- PMCID: PMC3740009
- DOI: 10.1038/nbt.2601
Network link prediction by global silencing of indirect correlations
Abstract
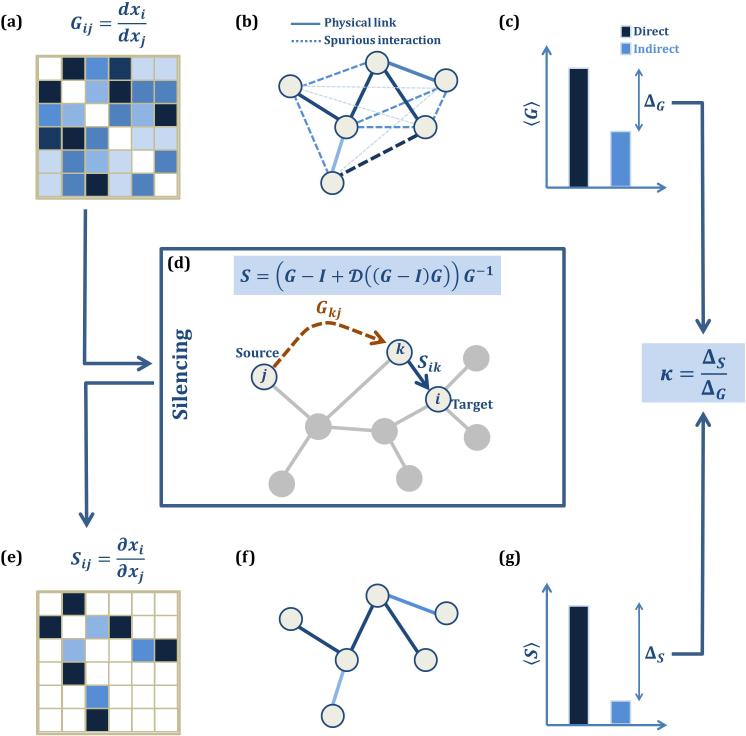
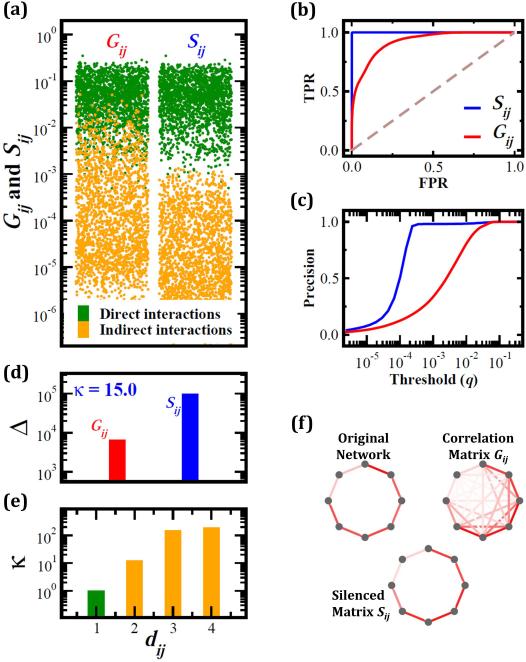
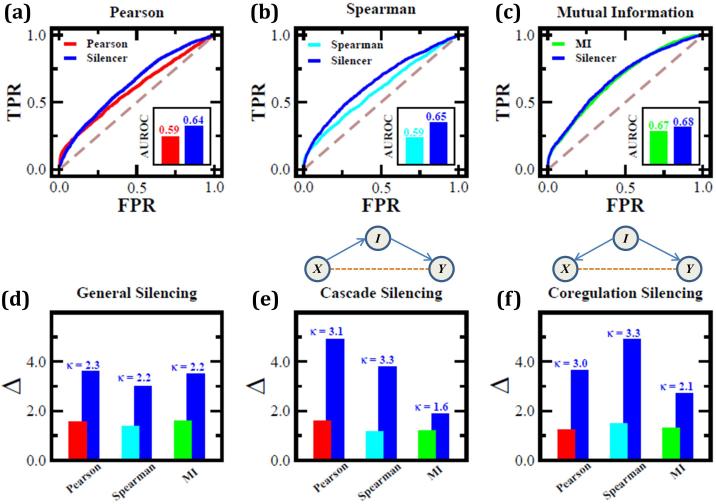
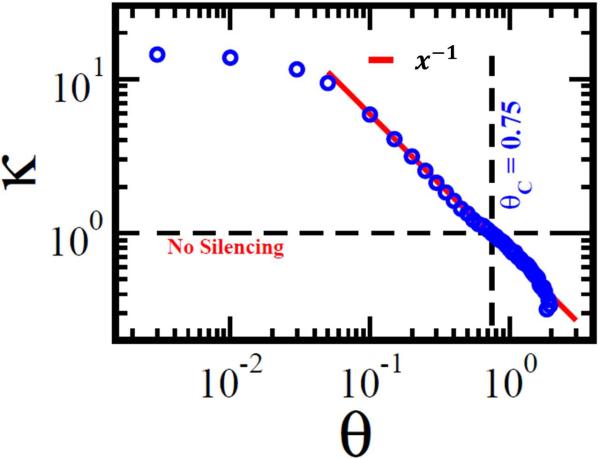
Predictions of physical and functional links between cellular components are often based on correlations between experimental measurements, such as gene expression. However, correlations are affected by both direct and indirect paths, confounding our ability to identify true pairwise interactions. Here we exploit the fundamental properties of dynamical correlations in networks to develop a method to silence indirect effects. The method receives as input the observed correlations between node pairs and uses a matrix transformation to turn the correlation matrix into a highly discriminative silenced matrix, which enhances only the terms associated with direct causal links. Against empirical data for Escherichia coli regulatory interactions, the method enhanced the discriminative power of the correlations by twofold, yielding >50% predictive improvement over traditional correlation measures and 6% over mutual information. Overall this silencing method will help translate the abundant correlation data into insights about a system's interactions, with applications ranging from link prediction to inferring the dynamical mechanisms governing biological networks.
Figures
Comment in
-
Network cleanup.Nat Biotechnol. 2013 Aug;31(8):714-5. doi: 10.1038/nbt.2657. Nat Biotechnol. 2013. PMID: 23929347 No abstract available.
-
Silence on the relevant literature and errors in implementation.Nat Biotechnol. 2015 Apr;33(4):336-9. doi: 10.1038/nbt.3185. Nat Biotechnol. 2015. PMID: 25850052 No abstract available.
-
Response to letter of correspondence - Bastiaens et al.Nat Biotechnol. 2015 Apr;33(4):339-42. doi: 10.1038/nbt.3184. Nat Biotechnol. 2015. PMID: 25850053 No abstract available.
References
-
- Vendruscolo M. In: Networks in Cell Biology. Buchanan M, Caldarelli G, De Los Rios P, Rao F, editors. Cambridge University Press; 2010.
-
- Kann MG. Protein interactions and disease: Computational approaches to uncover the etiology of diseases. Briefings in Bioinformatics. 2007;8:333–346. - PubMed
-
- Albert R. Scale-free networks in cell biology. Journal of Cell Science. 2005;118:4947–57. - PubMed
-
- Barabási A-L, Oltvai ZN. Network biology: understanding the cell's functional organization. Nature Reviews Genetics. 2004;5:101–113. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
