

Apomixis in plant reproduction: a novel perspective on an old dilemma
- PMID: 23852378
- PMCID: PMC3747320
- DOI: 10.1007/s00497-013-0222-y
Apomixis in plant reproduction: a novel perspective on an old dilemma
Abstract
Seed is one of the key factors of crop productivity. Therefore, a comprehension of the mechanisms underlying seed formation in cultivated plants is crucial for the quantitative and qualitative progress of agricultural production. In angiosperms, two pathways of reproduction through seed exist: sexual or amphimictic, and asexual or apomictic; the former is largely exploited by seed companies for breeding new varieties, whereas the latter is receiving continuously increasing attention from both scientific and industrial sectors in basic research projects. If apomixis is engineered into sexual crops in a controlled manner, its impact on agriculture will be broad and profound. In fact, apomixis will allow clonal seed production and thus enable efficient and consistent yields of high-quality seeds, fruits, and vegetables at lower costs. The development of apomixis technology is expected to have a revolutionary impact on agricultural and food production by reducing cost and breeding time, and avoiding the complications that are typical of sexual reproduction (e.g., incompatibility barriers) and vegetative propagation (e.g., viral transfer). However, the development of apomixis technology in agriculture requires a deeper knowledge of the mechanisms that regulate reproductive development in plants. This knowledge is a necessary prerequisite to understanding the genetic control of the apomictic process and its deviations from the sexual process. Our molecular understanding of apomixis will be greatly advanced when genes that are specifically or differentially expressed during embryo and embryo sac formation are discovered. In our review, we report the main findings on this subject by examining two approaches: i) analysis of the apomictic process in natural apomictic species to search for genes controlling apomixis and ii) analysis of gene mutations resembling apomixis or its components in species that normally reproduce sexually. In fact, our opinion is that a novel perspective on this old dilemma pertaining to the molecular control of apomixis can emerge from a cross-check among candidate genes in natural apomicts and a high-throughput analysis of sexual mutants.
Figures
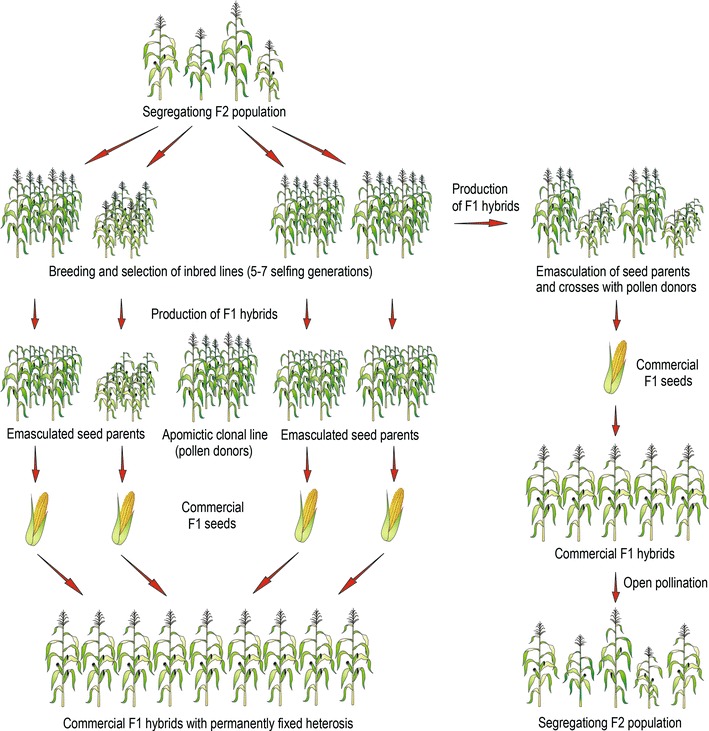
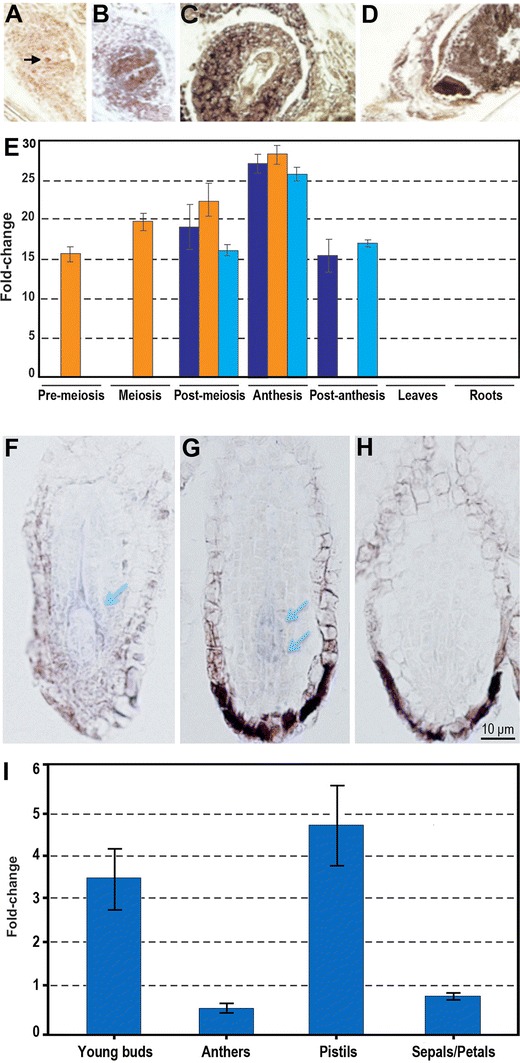
References
-
- Akiyama Y, Conner JA, Goel S, Morishige DT, Mullet JE, Hanna WW, Ozias-Akins P. High-resolution physical mapping in Pennisetum squamulatum reveals extensive chromosomal heteromorphism of the genomic region associated with apomixis. Plant Physiol. 2004;134:1733–1741. doi: 10.1104/pp.103.033969. - DOI - PMC - PubMed
-
- Albertini E, Barcaccia G (2007) Apomeiosis in Poa pratensis L. and Medicago sativa L. In: Horandl E, Grossniklaus U, Sharbel T, van Dijk P (eds) Apomixis: evolution, mechanisms and perspectives. Gantner, Ruggel [Regnum Veg. 147], pp 93–116
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
