

Inhibition of CD8+ T cell-derived CD40 signals is necessary but not sufficient for Foxp3+ induced regulatory T cell generation in vivo
- PMID: 23858029
- PMCID: PMC3735715
- DOI: 10.4049/jimmunol.1300267
Inhibition of CD8+ T cell-derived CD40 signals is necessary but not sufficient for Foxp3+ induced regulatory T cell generation in vivo
Abstract
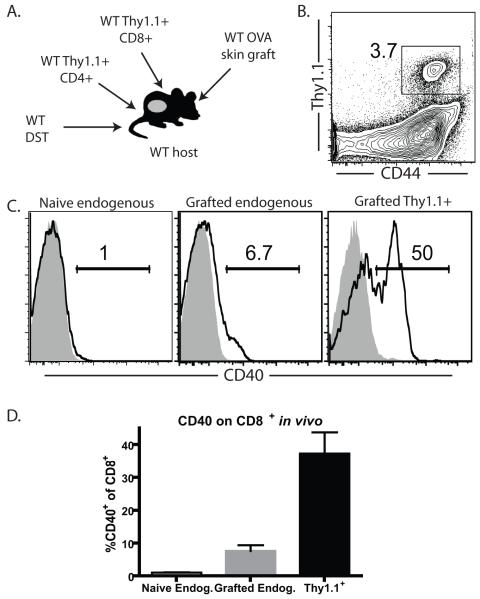
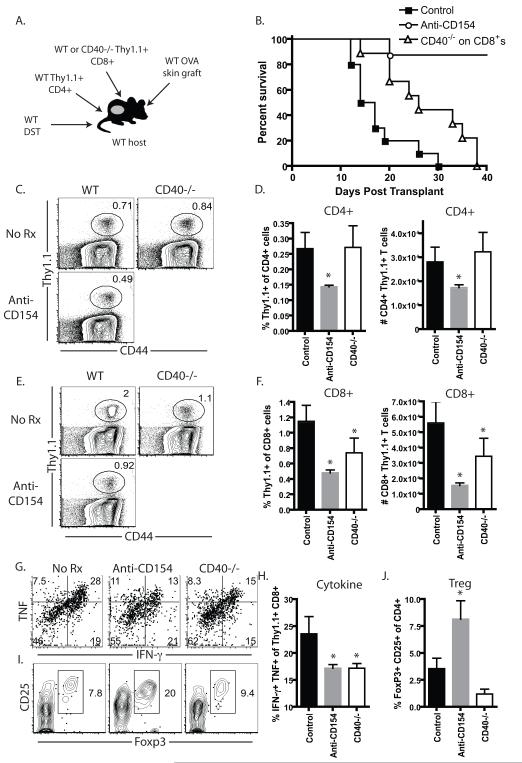
Current models of CD4(+) T cell help suggest a major role for CD154 binding to CD40 expressed on dendritic cells, with a lesser role for direct T:T interactions via CD40 expressed on CD8(+) T cells. However, the contribution of CD8(+) T cell-derived CD40 signals during the donor-reactive T cell response to a transplant has never been studied. In this study, we examined the graft-rejection kinetics and CD4(+) and CD8(+) donor-reactive T cell responses under conditions in which CD40 was genetically ablated only on APC, as well as under conditions in which CD40 was genetically ablated only on donor-reactive CD8(+) T cells. Our results revealed a significant role for CD8(+) T cell-expressed CD40 in the augmentation of donor-reactive CD8(+) T cell responses following transplantation and showed that CD40 expressed on CD8(+) T cells must be inhibited to allow conversion of CD4(+) T cells into induced regulatory T cells. Thus, this study identifies a major role for CD8(+) T cell-derived CD40 signals as a critical switch factor that both promotes optimal differentiation of cytokine-producing CD8(+) effector T cell responses and inhibits the differentiation of Ag-specific Foxp3(+) induced regulatory T cells in vivo.
Figures




References
-
- Bennett SRM, Carbone FR, Karamalis F, Flavell RA, Miller JFAP, Heath WR. Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature. 1998;393:478–480. - PubMed
-
- Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 1998;393:480–483. - PubMed
-
- Ridge JP, Di Rosa F, Matzinger P. A conditioned dendritic cell can be a temporal bridge between CD4+ T-helper and a T-killer cell. Nature. 1998;393:474–478. - PubMed
-
- Quezada SA, Jarvinen LZ, Lind EF, Noelle RJ. CD40/CD154 interactions at the interface of tolerance and immunity. Annu Rev Immunol. 2004;22:307–328. - PubMed
-
- Toes RE, Schoenberger SP, van der Voort EI, Offringa R, Melief CJ. CD40-CD40Ligand interactions and their role in cytotoxic T lymphocyte priming and anti-tumor immunity. Semin Immunol. 1998;10:443–448. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
