

B cell-specific deficiencies in mTOR limit humoral immune responses
- PMID: 23858034
- PMCID: PMC3906844
- DOI: 10.4049/jimmunol.1201767
B cell-specific deficiencies in mTOR limit humoral immune responses
Abstract
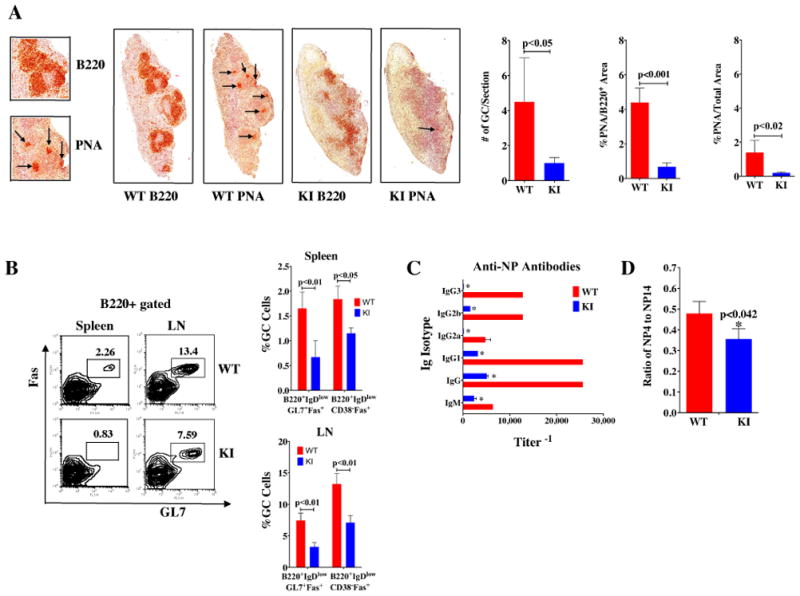
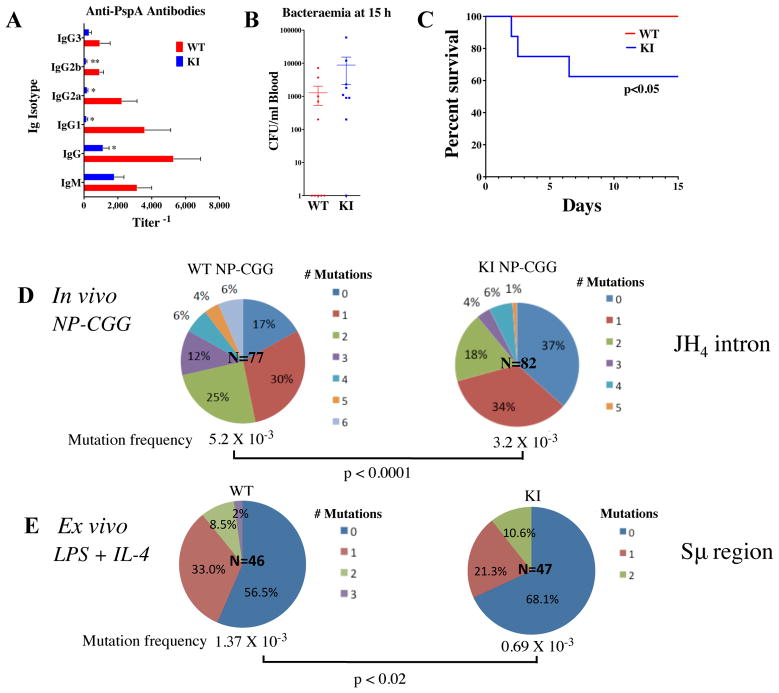
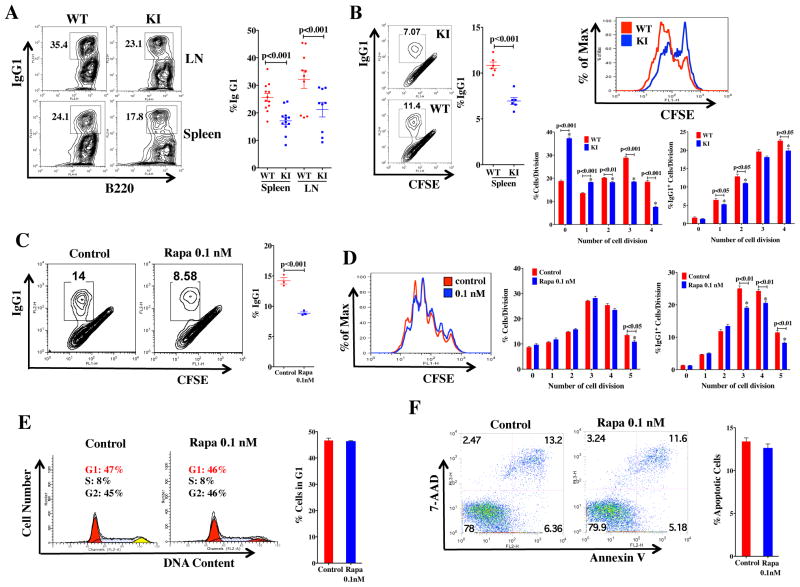
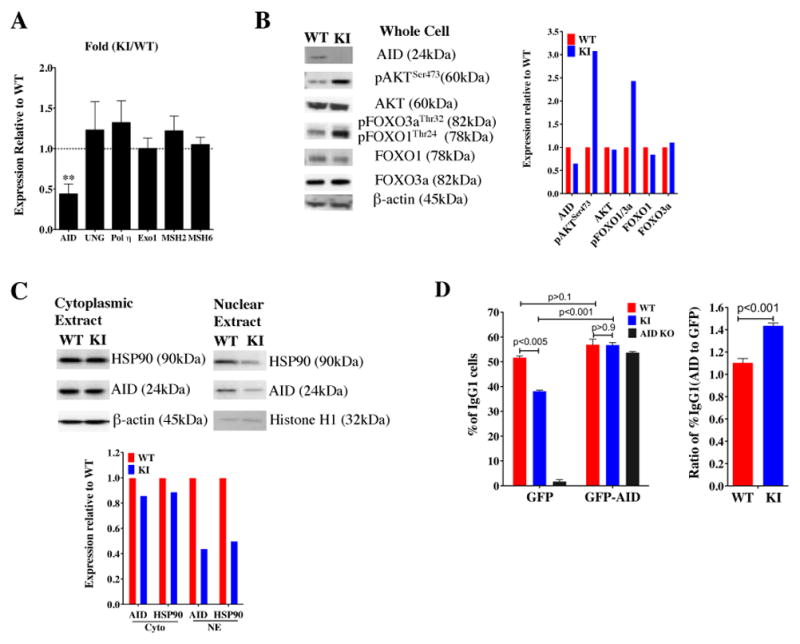
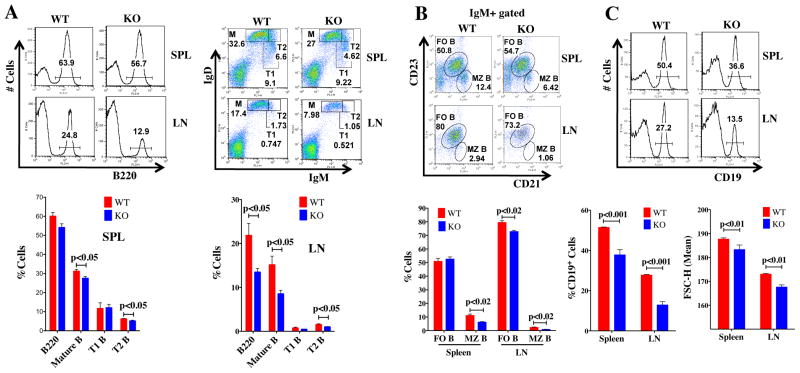
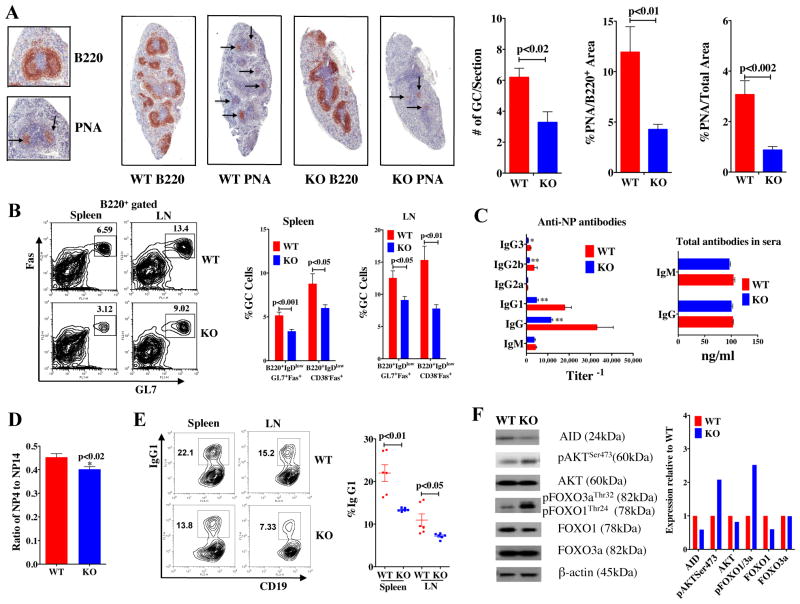
Generation of high-affinity Abs in response to Ags/infectious agents is essential for developing long-lasting immune responses. B cell maturation and Ab responses to Ag stimulation require Ig somatic hypermutation (SHM) and class-switch recombination (CSR) for high-affinity responses. Upon immunization with either the model Ag 4-hydroxy-3-nitrophenylacetyl hapten (NP) conjugated to chicken γ globulin lysine (NP-CGG) or heat-killed Streptococcus pneumoniae capsular type 14 protein (Pn14), knock-in (KI) mice hypomorphic for mTOR function had a decreased ability to form germinal centers, develop high-affinity anti-NP-specific or anti-Pn14-specific Abs, and perform SHM/CSR. Hypomorphic mTOR mice also had a high mortality (40%) compared with wild-type (WT) (0%) littermates and had lower pneumococcal surface protein A-specific Ab titers when immunized and challenged with live S. pneumoniae infection. Mice with mTOR deleted in their B cell lineage (knockout [KO]) also produced fewer splenic germinal centers and decreased high-affinity Ab responses to NP-CGG than did their WT littermates. CSR rates were lower in mTOR KI and KO mice, and pharmacologic inhibition of mTOR in WT B cells resulted in decreased rates of ex vivo CSR. RNA and protein levels of activation-induced cytidine deaminase (AID), a protein essential for SHM and CSR, were lower in B cells from both KI and B cell-specific KO mice, concomitant with increases in phosphorylated AKT and FOXO1. Rescue experiments increasing AID expression in KI B cells restored CSR levels to those in WT B cells. Thus, mTOR plays an important immunoregulatory role in the germinal center, at least partially through AID signaling, in generating high-affinity Abs.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
