

Talins and kindlins: partners in integrin-mediated adhesion
- PMID: 23860236
- PMCID: PMC4116690
- DOI: 10.1038/nrm3624
Talins and kindlins: partners in integrin-mediated adhesion
Abstract
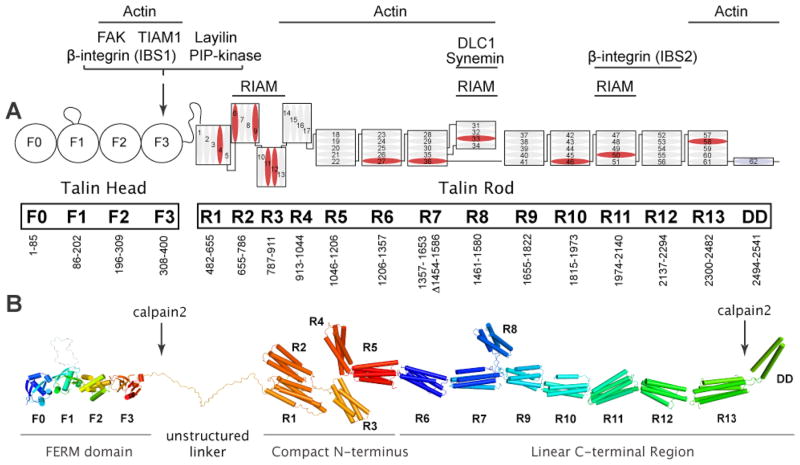
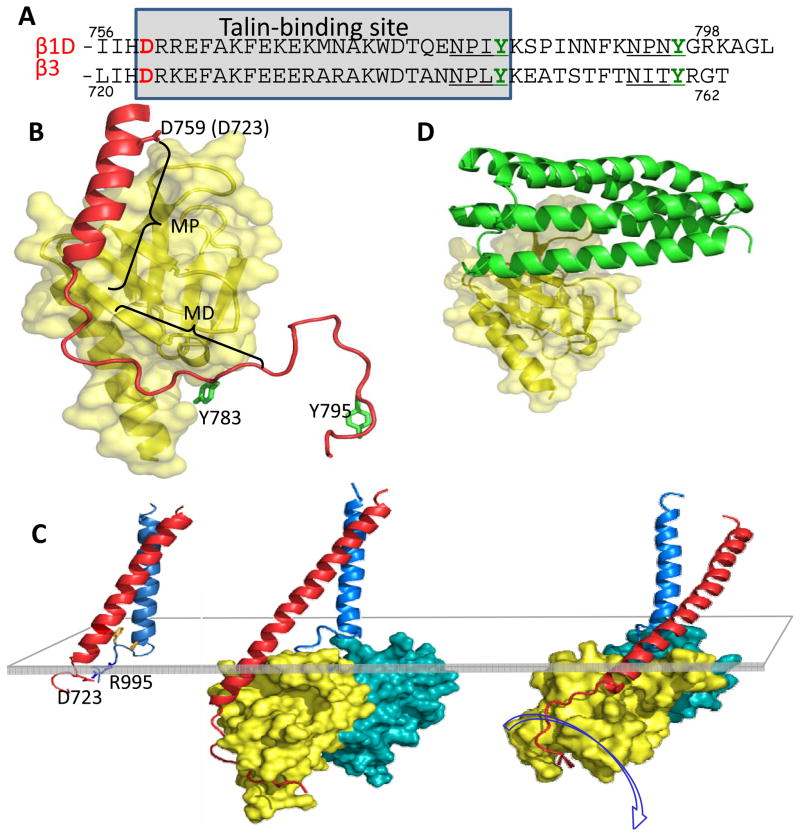
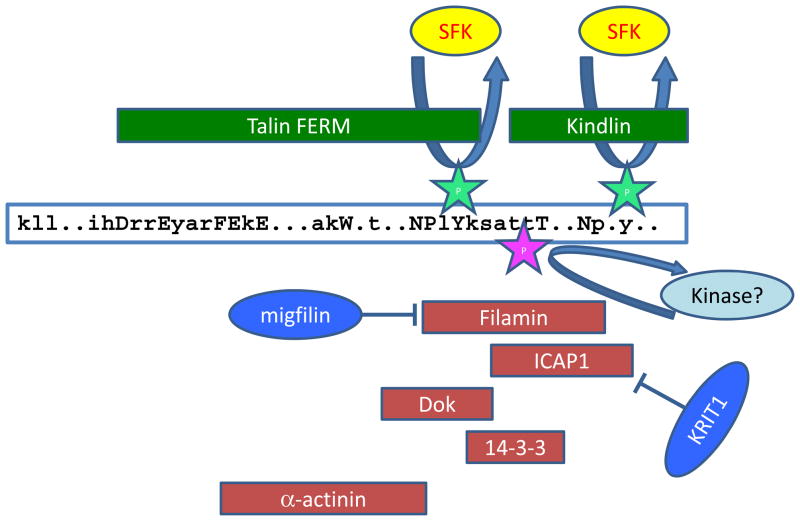
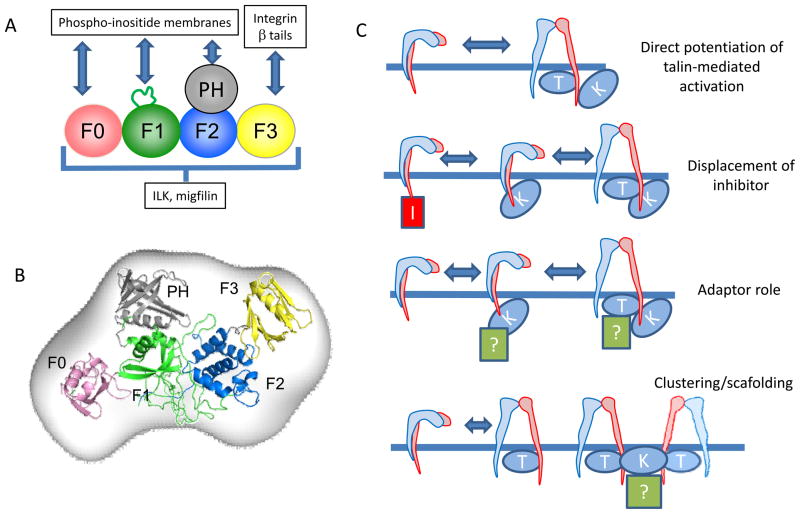
Integrin receptors provide a dynamic, tightly-regulated link between the extracellular matrix (or cellular counter-receptors) and intracellular cytoskeletal and signalling networks, enabling cells to sense and respond to their chemical and physical environment. Talins and kindlins, two families of FERM-domain proteins, bind the cytoplasmic tail of integrins, recruit cytoskeletal and signalling proteins involved in mechanotransduction and synergize to activate integrin binding to extracellular ligands. New data reveal the domain structure of full-length talin, provide insights into talin-mediated integrin activation and show that RIAM recruits talin to the plasma membrane, whereas vinculin stabilizes talin in cell-matrix junctions. How kindlins act is less well-defined, but disease-causing mutations show that kindlins are also essential for integrin activation, adhesion, cell spreading and signalling.
Figures
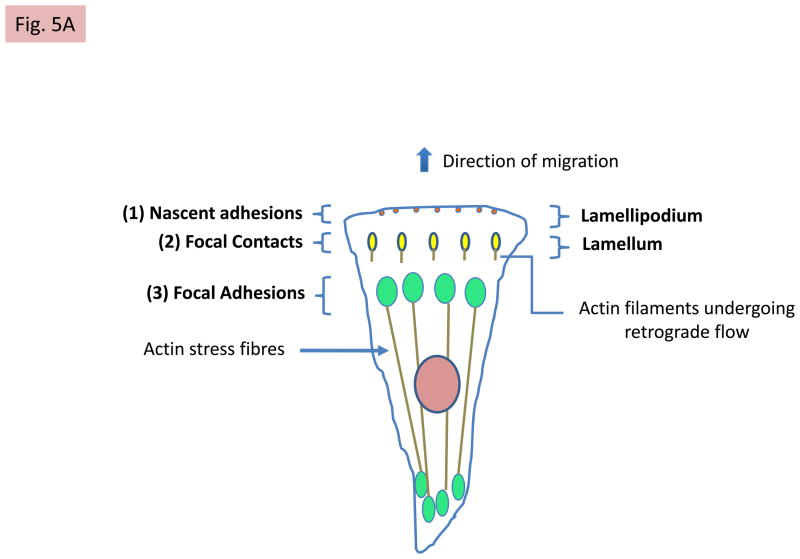
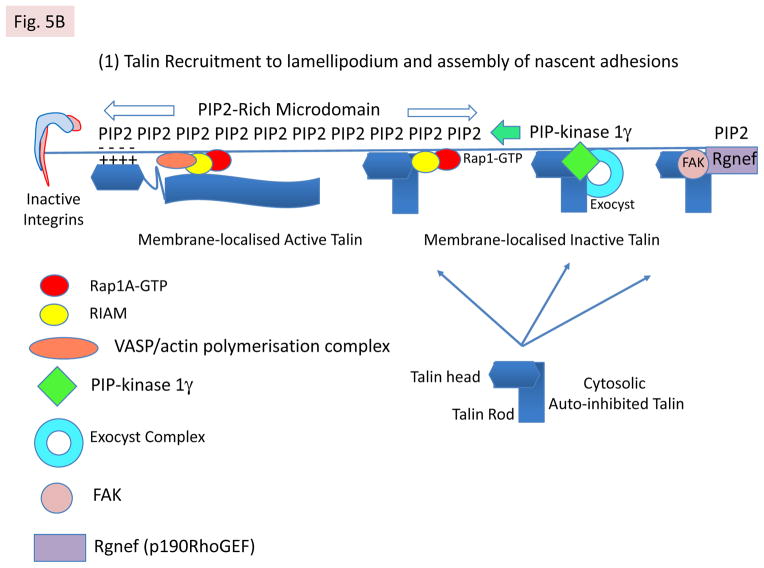
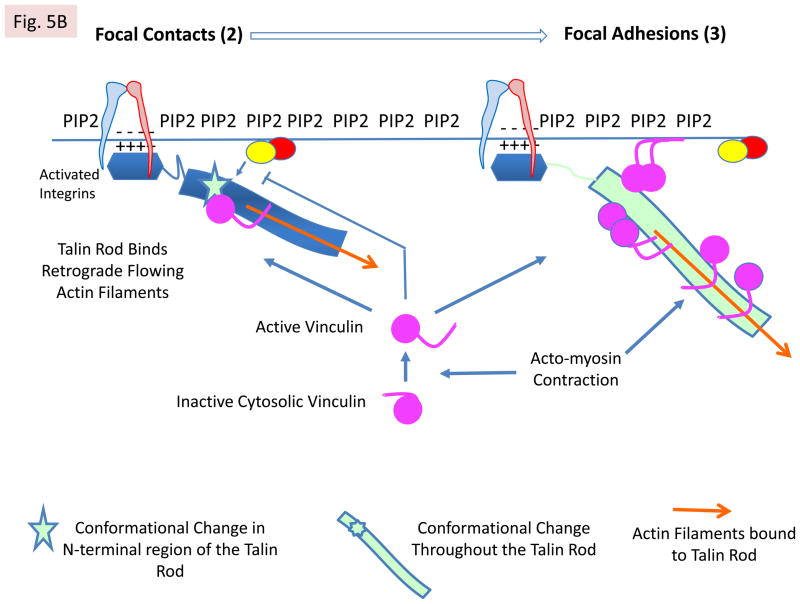
References
-
- Critchley DR. Biochemical and structural properties of the integrin-associated cytoskeletal protein talin. Annu Rev Biophys. 2009;38:235–54. - PubMed
-
- Calderwood DA, et al. The talin head domain binds to integrin b subunit cytoplasmic tails and regulates integrin activation. J Biol Chem. 1999;274:28071–28704. - PubMed
-
- Tadokoro S, et al. Talin binding to integrin beta tails: a final common step in integrin activation. Science. 2003;302:103–6. - PubMed
-
- Garcia-Alvarez B, et al. Structural determinants of integrin recognition by talin. Mol Cell. 2003;11:49–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
