

Brown fat in a protoendothermic mammal fuels eutherian evolution
- PMID: 23860571
- PMCID: PMC3717499
- DOI: 10.1038/ncomms3140
Brown fat in a protoendothermic mammal fuels eutherian evolution
Abstract
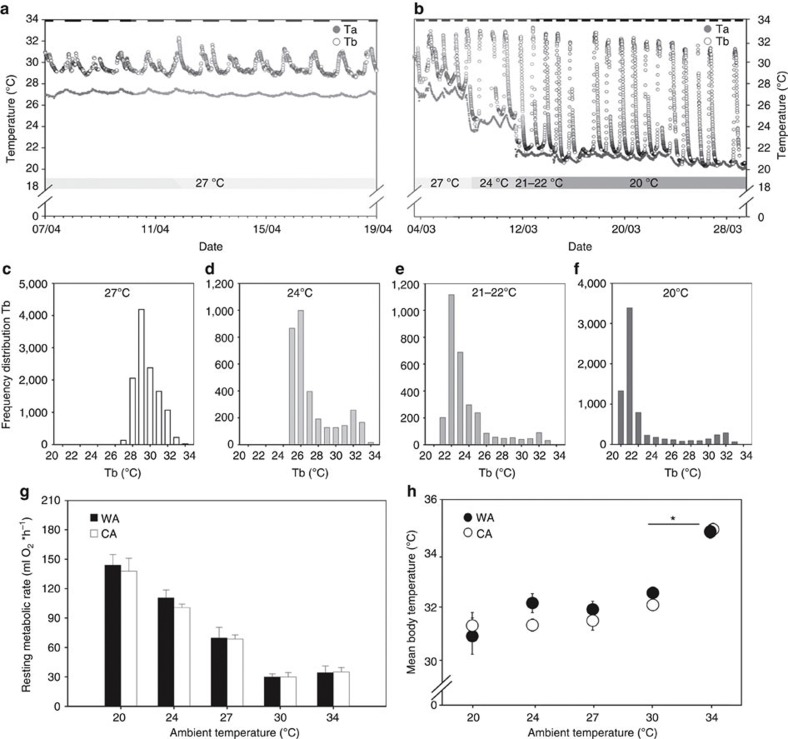
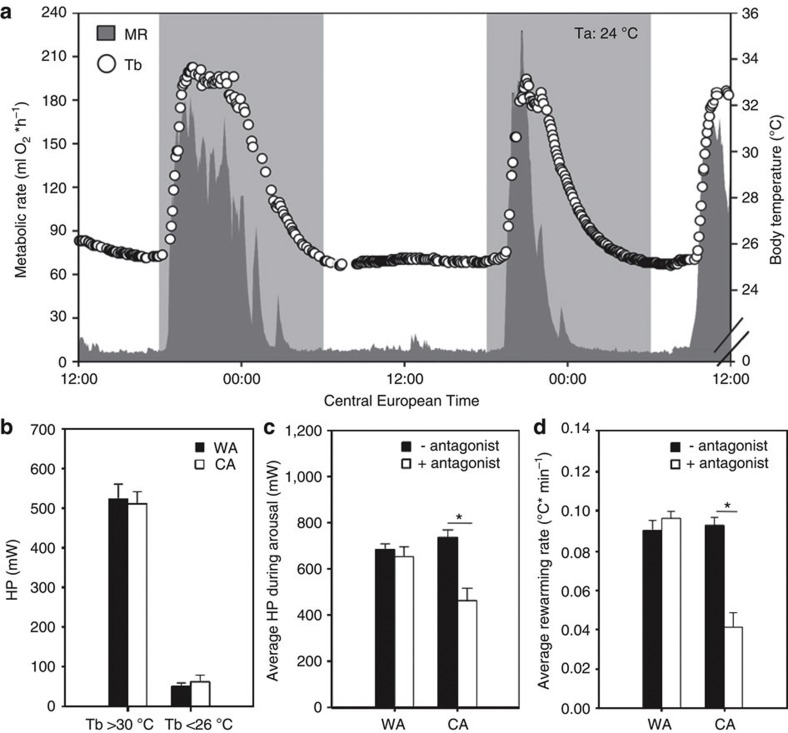
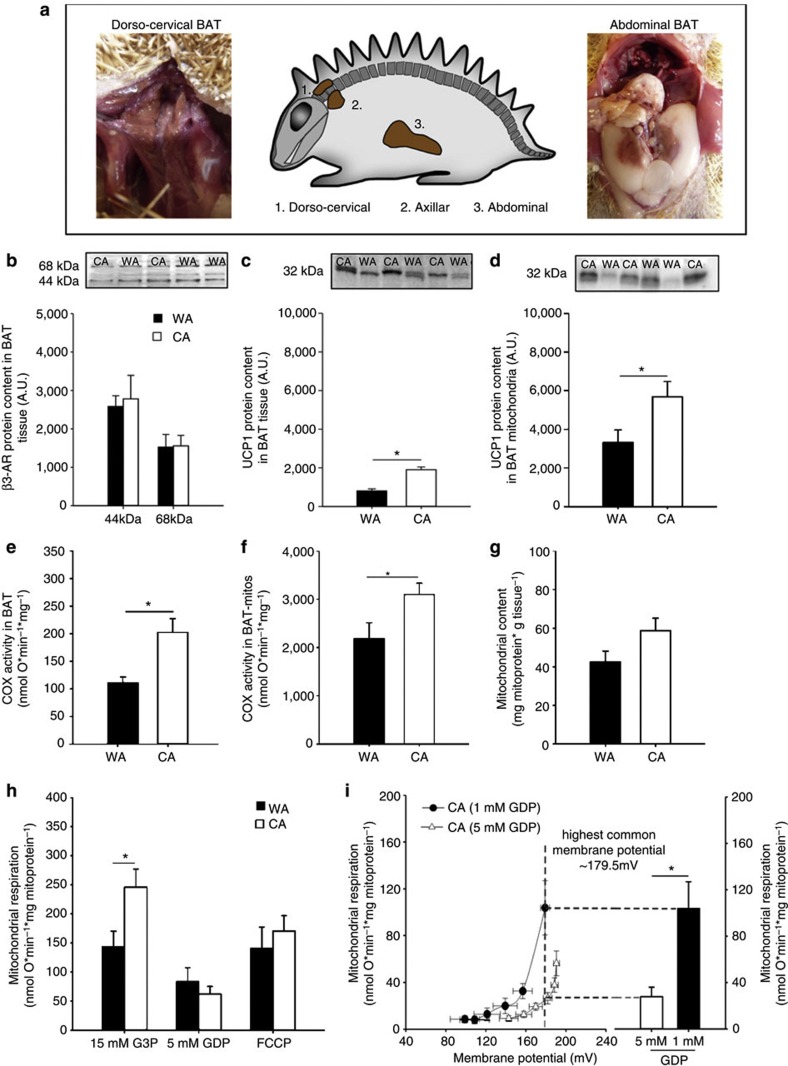
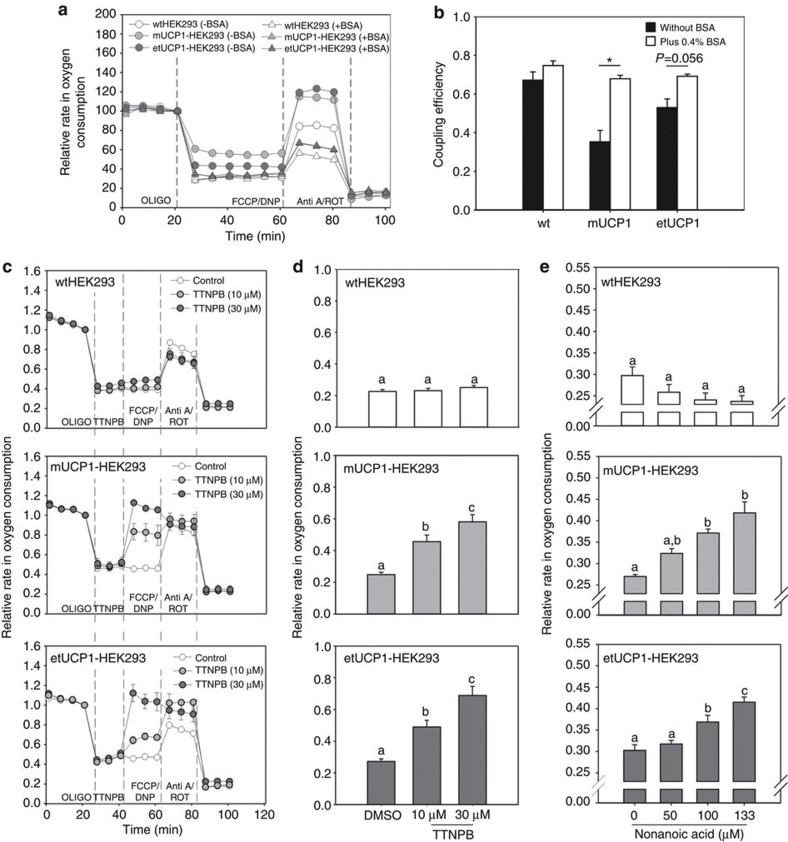
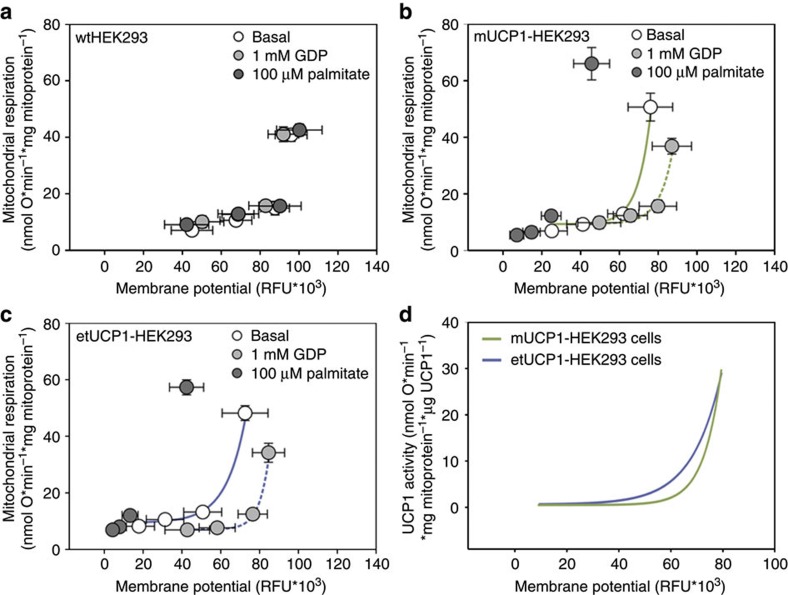
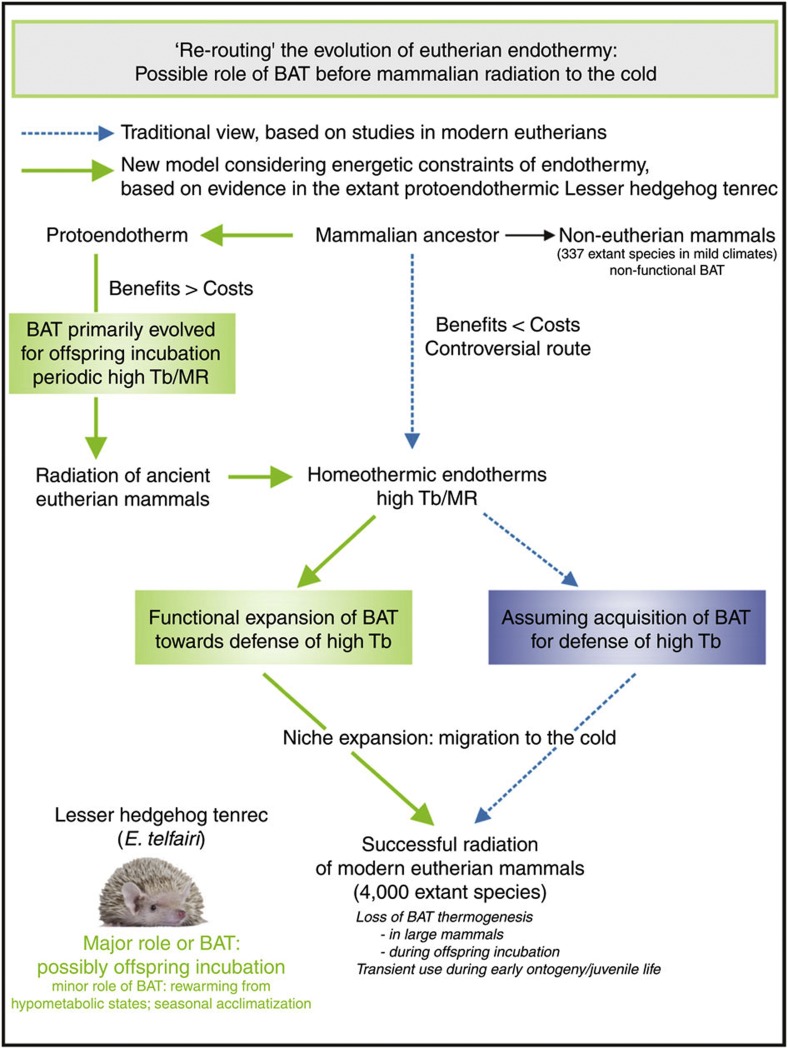
Endothermy has facilitated mammalian species radiation, but the sequence of events leading to sustained thermogenesis is debated in multiple evolutionary models. Here we study the Lesser hedgehog tenrec (Echinops telfairi), a phylogenetically ancient, 'protoendothermic' eutherian mammal, in which constantly high body temperatures are reported only during reproduction. Evidence for nonshivering thermogenesis is found in vivo during periodic ectothermic-endothermic transitions. Anatomical studies reveal large brown fat-like structures in the proximity of the reproductive organs, suggesting physiological significance for parental care. Biochemical analysis demonstrates high mitochondrial proton leak catalysed by an uncoupling protein 1 ortholog. Strikingly, bioenergetic profiling of tenrec uncoupling protein 1 reveals similar thermogenic potency as modern mouse uncoupling protein 1, despite the large phylogenetic distance. The discovery of functional brown adipose tissue in this 'protoendothermic' mammal links nonshivering thermogenesis directly to the roots of eutherian evolution, suggesting physiological importance prior to sustained body temperatures and migration to the cold.
Figures
References
-
- Crompton A. W., Taylor C. R. & Jagger J. A. Evolution of homoeothermy in mammals. Nature 272, 333–336 (1978). - PubMed
-
- Lovegrove B. G. The evolution of endothermy in Cenozoic mammals: a plesiomorphic- apomorphic continuum. Biol. Rev. Camb. Philos. Soc. 87, 128–162 (2012). - PubMed
-
- McNab B. K. The evolution of homoeothermy in the phylogeny of mammals. Amer. Nat. 112, 1–21 (1978).
-
- Farmer C. G. Parental care: the key to understanding endothermy and other convergent features in birds and mammals. Am. Nat. 155, 326–334 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
