

AMP-activated protein kinase regulates the vacuolar H+-ATPase via direct phosphorylation of the A subunit (ATP6V1A) in the kidney
- PMID: 23863464
- PMCID: PMC3798744
- DOI: 10.1152/ajprenal.00303.2013
AMP-activated protein kinase regulates the vacuolar H+-ATPase via direct phosphorylation of the A subunit (ATP6V1A) in the kidney
Abstract
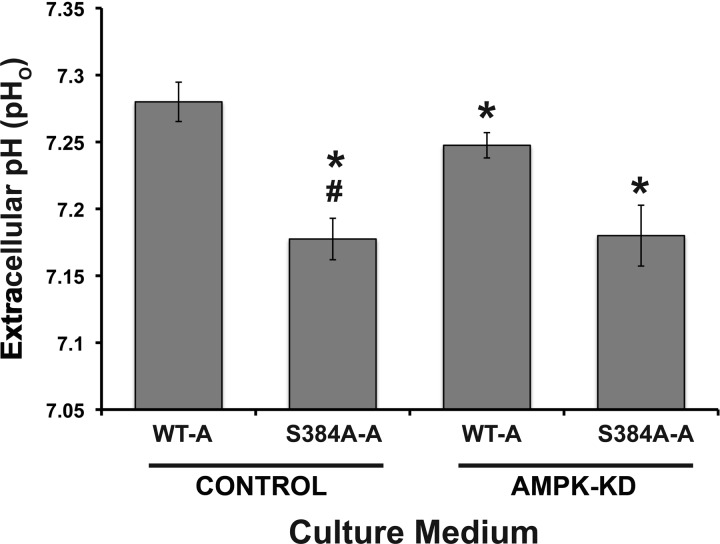
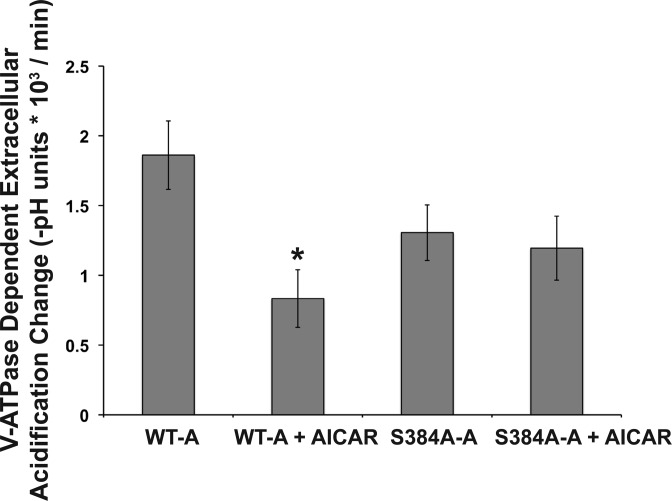
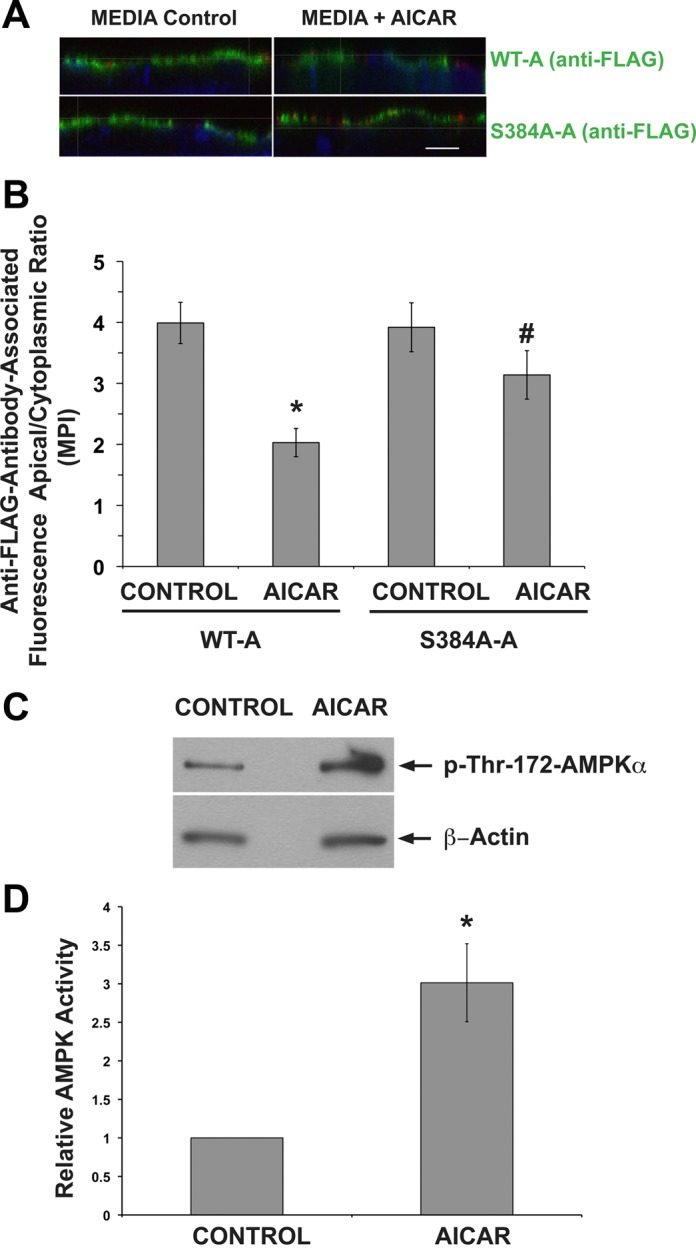
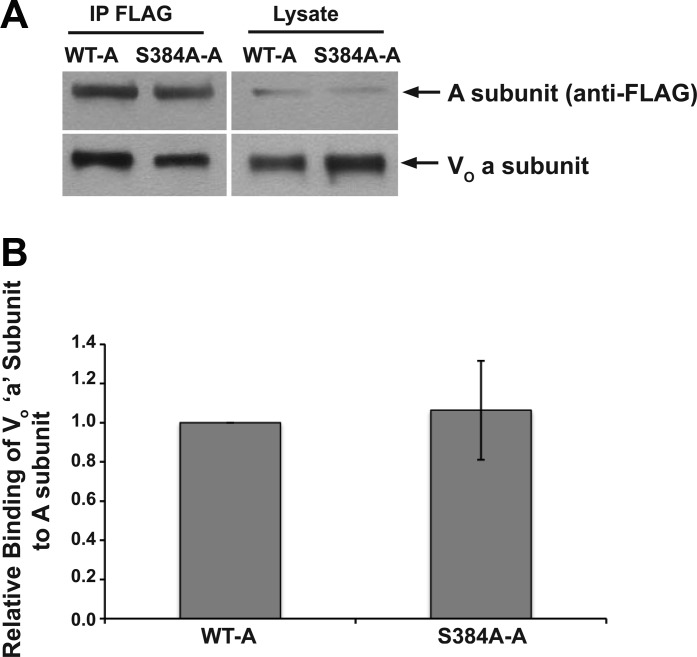
The vacuolar H(+)-ATPase (V-ATPase) in intercalated cells contributes to luminal acidification in the kidney collecting duct and nonvolatile acid excretion. We previously showed that the A subunit in the cytoplasmic V1 sector of the V-ATPase (ATP6V1A) is phosphorylated by the metabolic sensor AMP-activated protein kinase (AMPK) in vitro and in kidney cells. Here, we demonstrate that treatment of rabbit isolated, perfused collecting ducts with the AMPK activator 5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside (AICAR) inhibited V-ATPase-dependent H(+) secretion from intercalated cells after an acid load. We have identified by mass spectrometry that Ser-384 is a major AMPK phosphorylation site in the V-ATPase A subunit, a result confirmed by comparing AMPK-dependent phosphate labeling of wild-type A-subunit (WT-A) with that of a Ser-384-to-Ala A subunit mutant (S384A-A) in vitro and in intact HEK-293 cells. Compared with WT-A-expressing HEK-293 cells, S384A-A-expressing cells exhibited greater steady-state acidification of HCO3(-)-containing media. Moreover, AICAR treatment of clone C rabbit intercalated cells expressing the WT-A subunit reduced V-ATPase-dependent extracellular acidification, an effect that was blocked in cells expressing the phosphorylation-deficient S384A-A mutant. Finally, expression of the S384A-A mutant prevented cytoplasmic redistribution of the V-ATPase by AICAR in clone C cells. In summary, direct phosphorylation of the A subunit at Ser-384 by AMPK represents a novel regulatory mechanism of the V-ATPase in kidney intercalated cells. Regulation of the V-ATPase by AMPK may couple V-ATPase activity to cellular metabolic status with potential relevance to ischemic injury in the kidney and other tissues.
Keywords: AMPK; Intercalated cells; V-ATPase; kidney; mass spectrometry.
Figures
Comment in
-
Connecting type A intercalated cell metabolic state to V-ATPase function: phosphorylation does matter!Am J Physiol Renal Physiol. 2013 Oct 15;305(8):F1105-6. doi: 10.1152/ajprenal.00428.2013. Epub 2013 Jul 31. Am J Physiol Renal Physiol. 2013. PMID: 23904225 Free PMC article. No abstract available.
References
-
- Al-Awqati Q, Gao XB. Differentiation of intercalated cells in the kidney. Physiology (Bethesda) 26: 266–272, 2011 - PubMed
-
- Alzamora R, Thali RF, Gong F, Smolak C, Li H, Baty CJ, Bertrand CA, Auchli Y, Brunisholz RA, Neumann D, Hallows KR, Pastor-Soler NM. PKA regulates vacuolar H+-ATPase localization and activity via direct phosphorylation of the A subunit in kidney cells. J Biol Chem 285: 24676–24685, 2010 - PMC - PubMed
-
- Bens M, Vallet V, Cluzeaud F, Pascual-Letallec L, Kahn A, Rafestin-Oblin ME, Rossier BC, Vandewalle A. Corticosteroid-dependent sodium transport in a novel immortalized mouse collecting duct principal cell line. J Am Soc Nephrol 10: 923–934, 1999 - PubMed
-
- Bhalla V, Oyster NM, Fitch AC, Wijngaarden MA, Neumann D, Schlattner U, Pearce D, Hallows KR. AMP-activated kinase inhibits the epithelial Na+ channel through functional regulation of the ubiquitin ligase Nedd4–2. J Biol Chem 281: 26159–26169, 2006 - PubMed
-
- Bouley R, Pastor-Soler N, Cohen O, McLaughlin M, Breton S, Brown D. Stimulation of AQP2 membrane insertion in renal epithelial cells in vitro and in vivo by the cGMP phosphodiesterase inhibitor sildenafil citrate (Viagra). Am J Physiol Renal Physiol 288: F1103–F1112, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous