

Persistent synaptic scaling independent of AMPA receptor subunit composition
- PMID: 23864664
- PMCID: PMC6794055
- DOI: 10.1523/JNEUROSCI.1102-13.2013
Persistent synaptic scaling independent of AMPA receptor subunit composition
Abstract
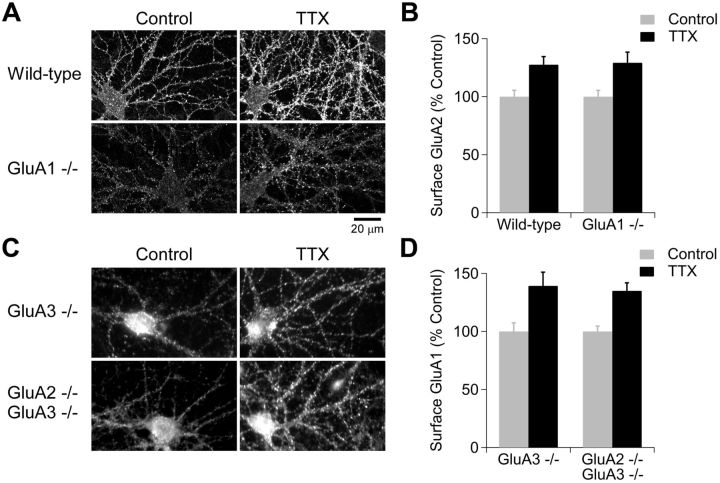
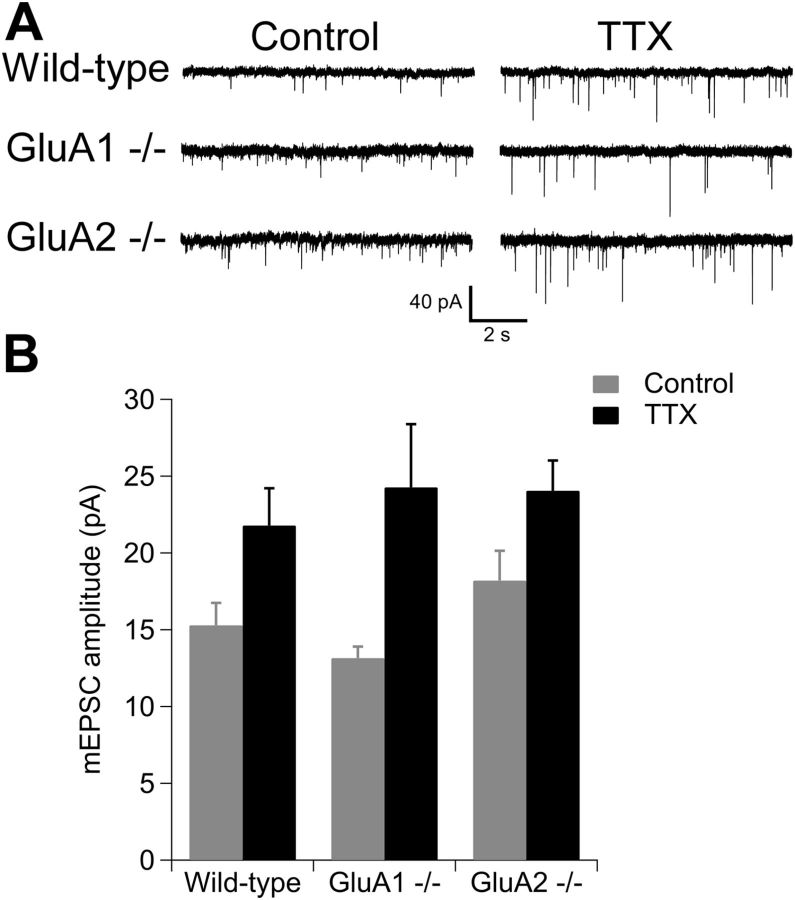
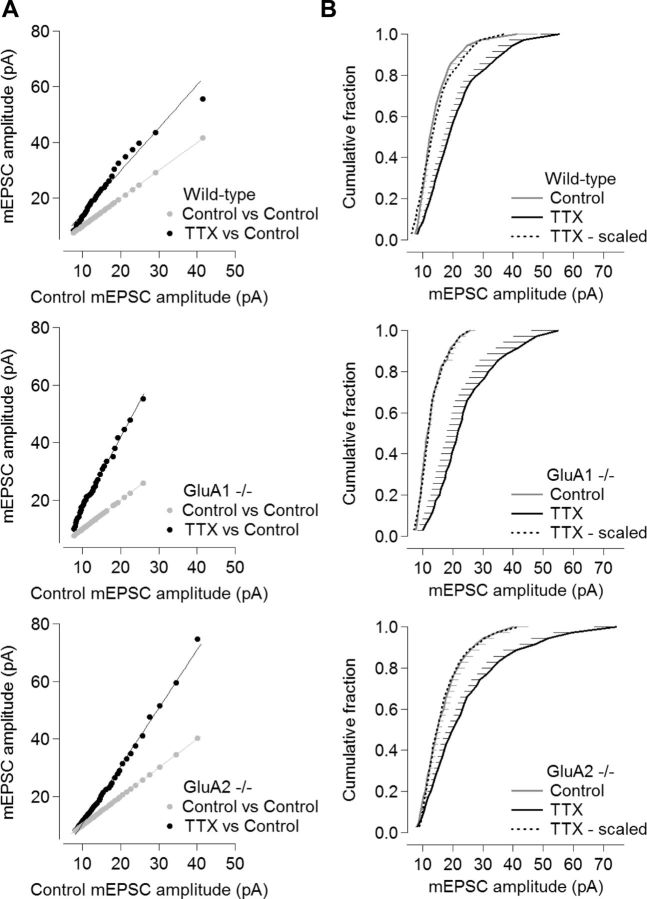
Despite long-standing evidence that the specific intracellular domains of AMPA-type glutamate receptor (AMPAR) subunits are critical for trafficking, it has recently been demonstrated that there is no absolute requirement for any AMPAR subunit for the receptor insertion underlying LTP. It is unclear whether this holds true to other forms of plasticity. Homeostatic synaptic plasticity (HSP) is an important form of negative feedback that provides stability to neuronal networks, and results at least in part from the insertion of AMPARs into glutamatergic synapses following chronic reductions in neuronal activity. Similar to LTP, the GluA1 subunit has been suggested to be the requisite subunit for HSP-induced AMPAR insertion and acute treatment with signaling molecules that underlie some forms of HSP results in the preferential incorporation of GluA2-lacking receptors. However, knockdown experiments have instead implicated a requirement for the GluA2 subunit. Here we re-examined the requirement for specific AMPAR subunit during chronic tetrodotoxin-induced HSP using hippocampal cultures derived from AMPAR subunit knock-out mice. We observed HSP in cultures from GluA1⁻/⁻, GluA2⁻/⁻, and GluA2⁻/⁻ GluA3⁻/⁻ mice, and conclude that, as with LTP, there is no subunit requirement for HSP.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials