

Calpain A modulates Toll responses by limited Cactus/IκB proteolysis
- PMID: 23864715
- PMCID: PMC3771957
- DOI: 10.1091/mbc.E13-02-0113
Calpain A modulates Toll responses by limited Cactus/IκB proteolysis
Abstract
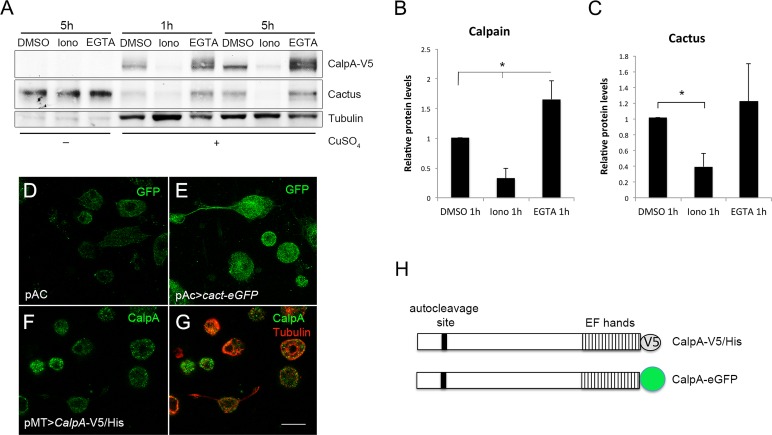
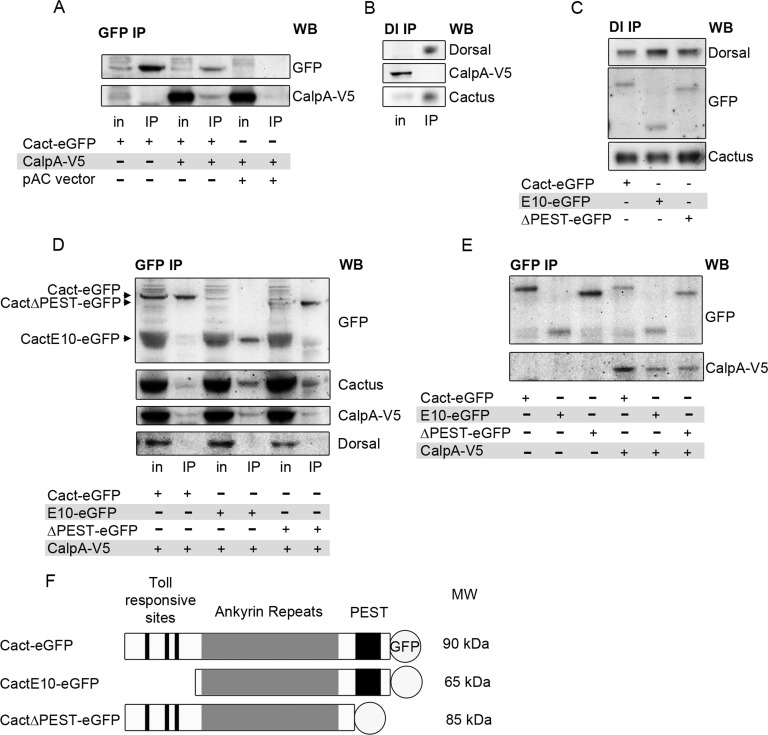
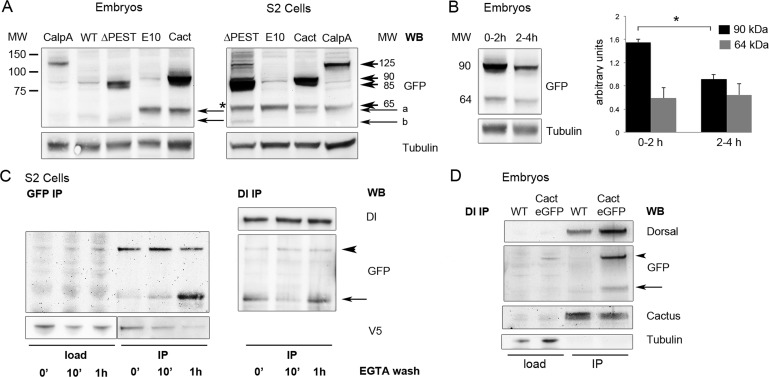
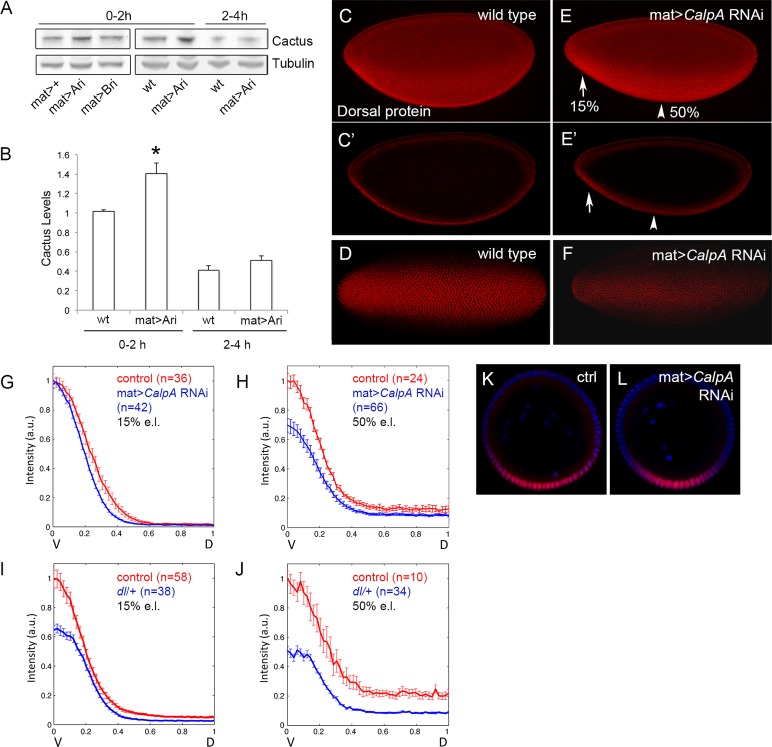
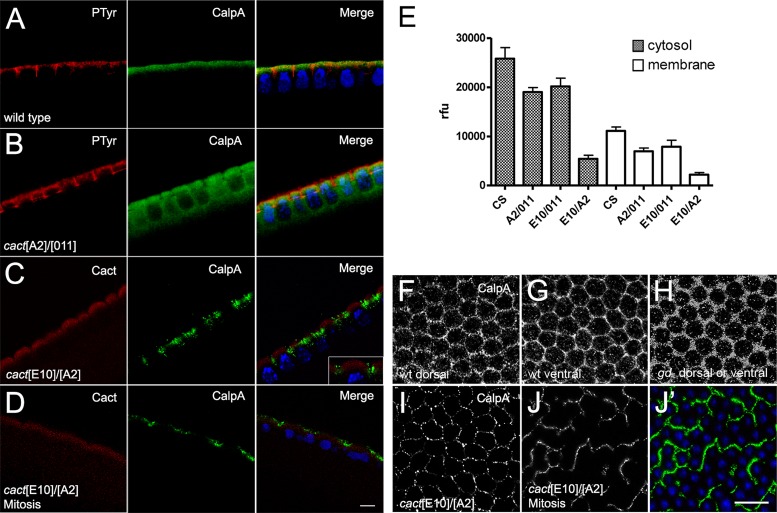
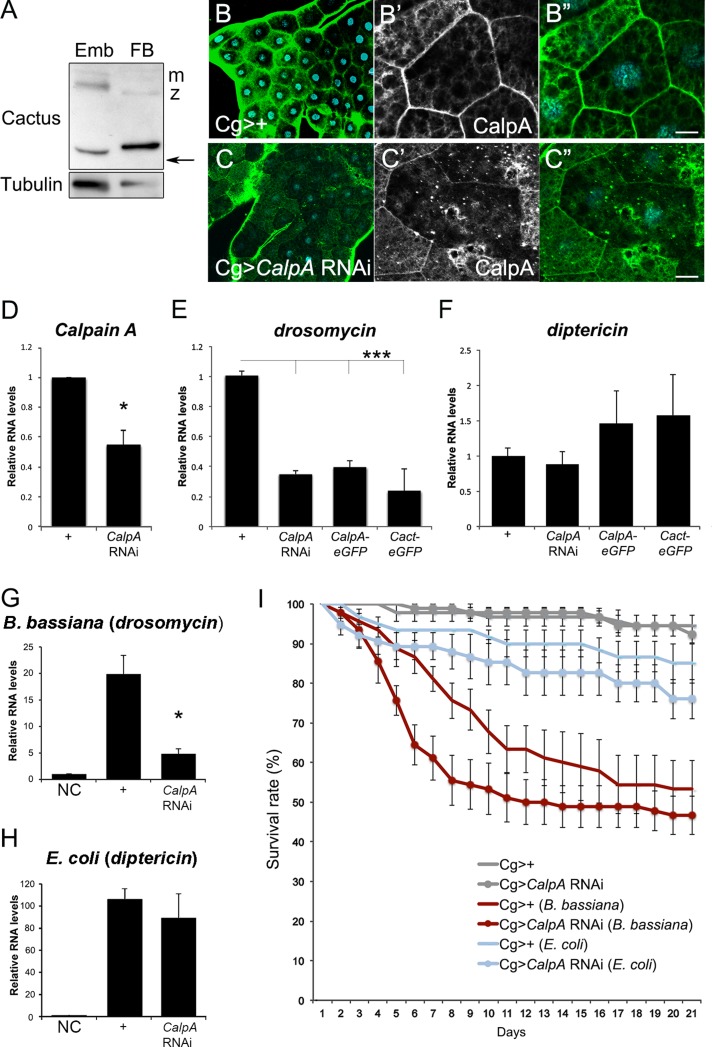
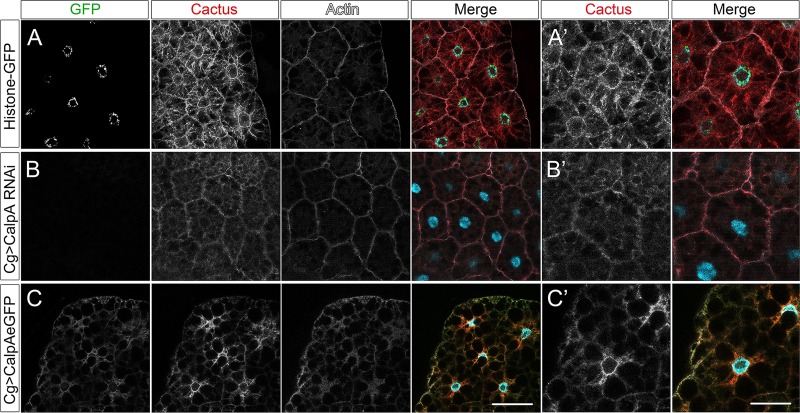
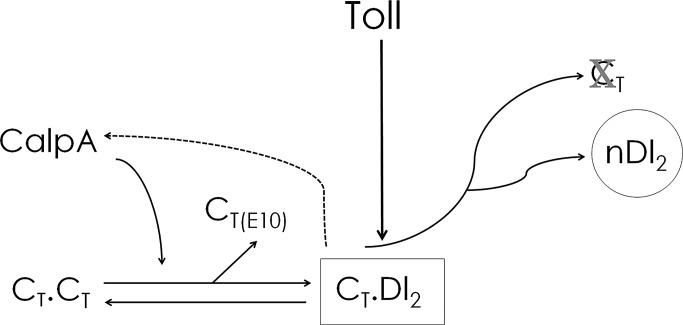
Calcium-dependent cysteine proteases of the calpain family are modulatory proteases that cleave their substrates in a limited manner. Among their substrates, calpains target vertebrate and invertebrate IκB proteins. Because proteolysis by calpains potentially generates novel protein functions, it is important to understand how this affects NFκB activity. We investigate the action of Calpain A (CalpA) on the Drosophila melanogaster IκB homologue Cactus in vivo. CalpA alters the absolute amounts of Cactus protein. Our data indicate, however, that CalpA uses additional mechanisms to regulate NFκB function. We provide evidence that CalpA interacts physically with Cactus, recognizing a Cactus pool that is not bound to Dorsal, a fly NFκB/Rel homologue. We show that proteolytic cleavage by CalpA generates Cactus fragments lacking an N-terminal region required for Toll responsiveness. These fragments are generated in vivo and display properties distinct from those of full-length Cactus. We propose that CalpA targets free Cactus, which is incorporated into and modulates Toll-responsive complexes in the embryo and immune system.
Figures
References
-
- Anderson KV, Jurgens G, Nusslein-Volhard C. Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell. 1985;42:779–789. - PubMed
-
- Araujo H, Negreiros E, Bier E. Integrins modulate Sog activity in the Drosophila wing. Development. 2003;130:3851–3864. - PubMed
-
- Baghdiguian S, et al. Calpain 3 deficiency is associated with myonuclear apoptosis and profound perturbation of the IkappaB alpha/NF-kappaB pathway in limb-girdle muscular dystrophy type 2A. Nat Med. 1999;5:503–511. - PubMed
-
- Baud V, Derudder E. Control of NF-kappaB activity by proteolysis. Curr Top Microbiol Immunol. 2011;349:97–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
