

Transplantation of human fetal-derived neural stem cells improves cognitive function following cranial irradiation
- PMID: 23866792
- PMCID: PMC3895108
- DOI: 10.3727/096368913X670200
Transplantation of human fetal-derived neural stem cells improves cognitive function following cranial irradiation
Abstract
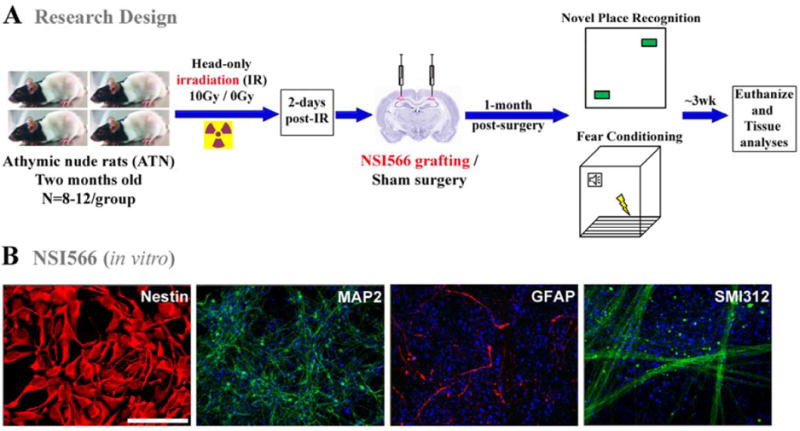
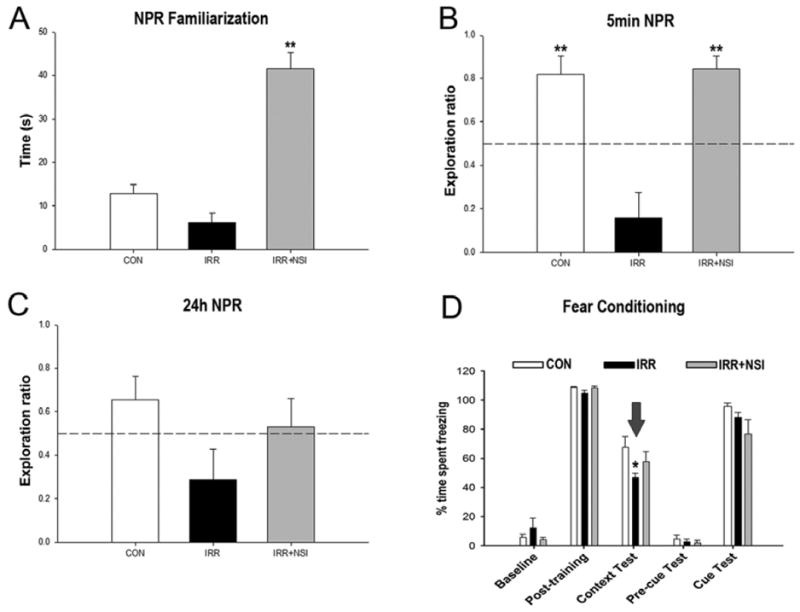
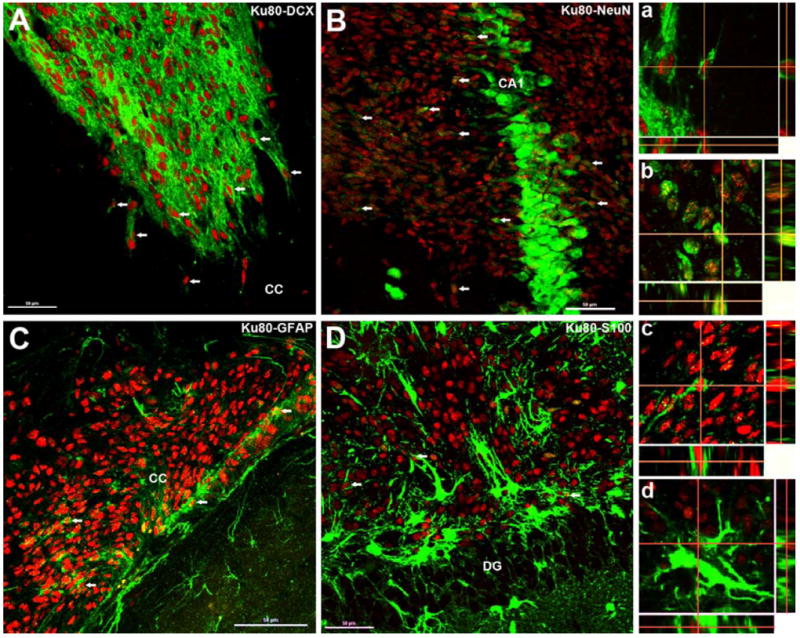
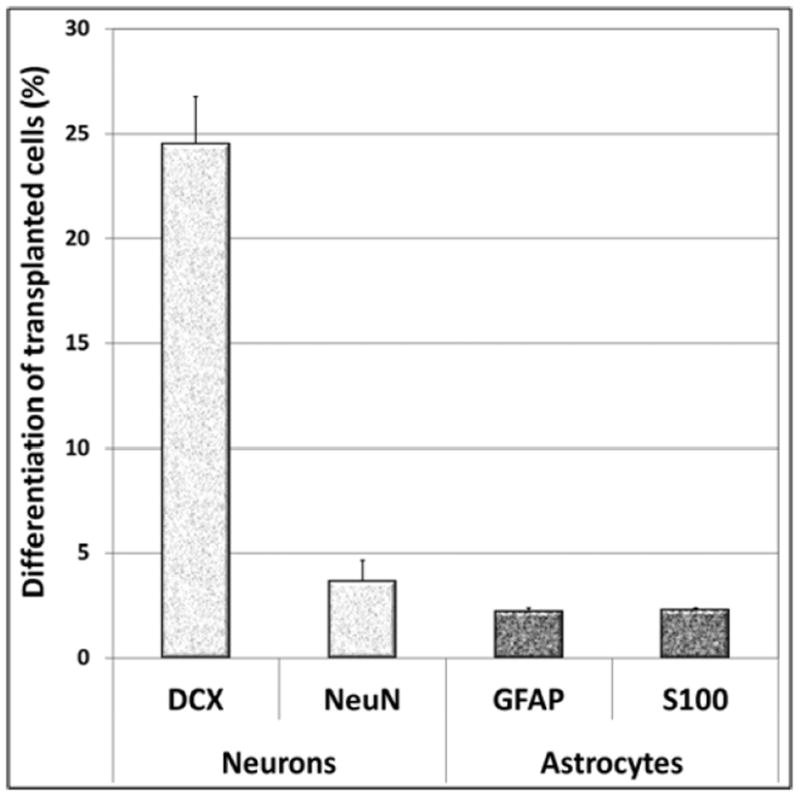
Treatment of central nervous system (CNS) malignancies typically involves radiotherapy to forestall tumor growth and recurrence following surgical resection. Despite the many benefits of cranial radiotherapy, survivors often suffer from a wide range of debilitating and progressive cognitive deficits. Thus, while patients afflicted with primary and secondary malignancies of the CNS now experience longer local regional control and progression-free survival, there remains no clinical recourse for the unintended neurocognitive sequelae associated with their cancer treatments. Multiple mechanisms contribute to disrupted cognition following irradiation, including the depletion of radiosensitive populations of stem and progenitor cells in the hippocampus. We have explored the potential of using intrahippocampal transplantation of human stem cells to ameliorate radiation-induced cognitive dysfunction. Past studies demonstrated the capability of cranially transplanted human embryonic (hESCs) and neural (hNSCs) stem cells to functionally restore cognition in rats 1 and 4 months after cranial irradiation. The present study employed an FDA-approved fetal-derived hNSC line capable of large scale-up under good manufacturing practice (GMP). Animals receiving cranial transplantation of these cells 1 month following irradiation showed improved hippocampal spatial memory and contextual fear conditioning performance compared to irradiated, sham surgery controls. Significant newly born (doublecortin positive) neurons and a smaller fraction of glial subtypes were observed within and nearby the transplantation core. Engrafted cells migrated and differentiated into neuronal and glial subtypes throughout the CA1 and CA3 subfields of the host hippocampus. These studies expand our prior findings to demonstrate that transplantation of fetal-derived hNSCs improves cognitive deficits in irradiated animals, as assessed by two separate cognitive tasks.
Conflict of interest statement
Authors declare no conflicts of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
