

Collagen VII plays a dual role in wound healing
- PMID: 23867500
- PMCID: PMC3726167
- DOI: 10.1172/JCI68127
Collagen VII plays a dual role in wound healing
Abstract
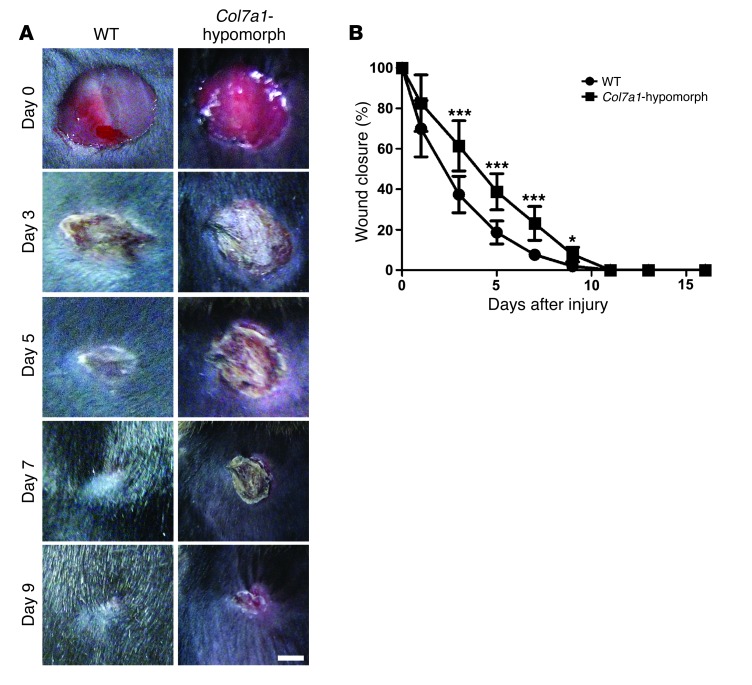
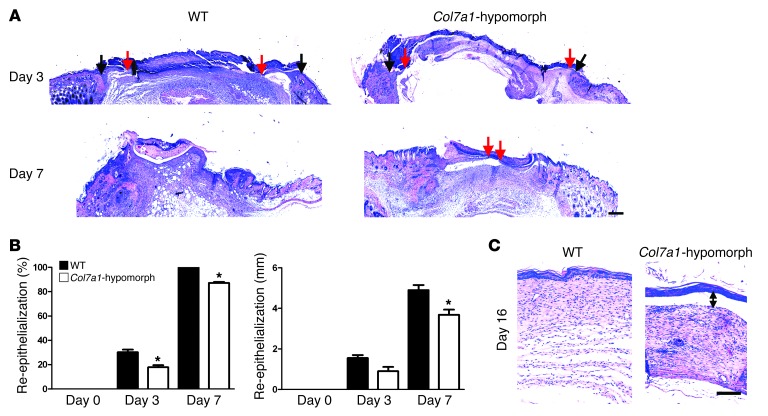
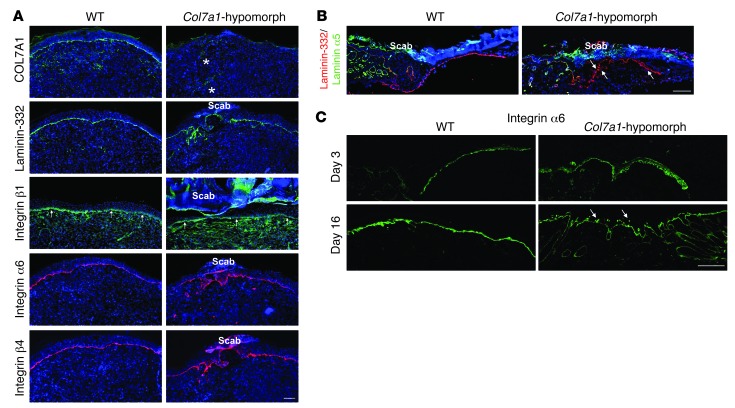
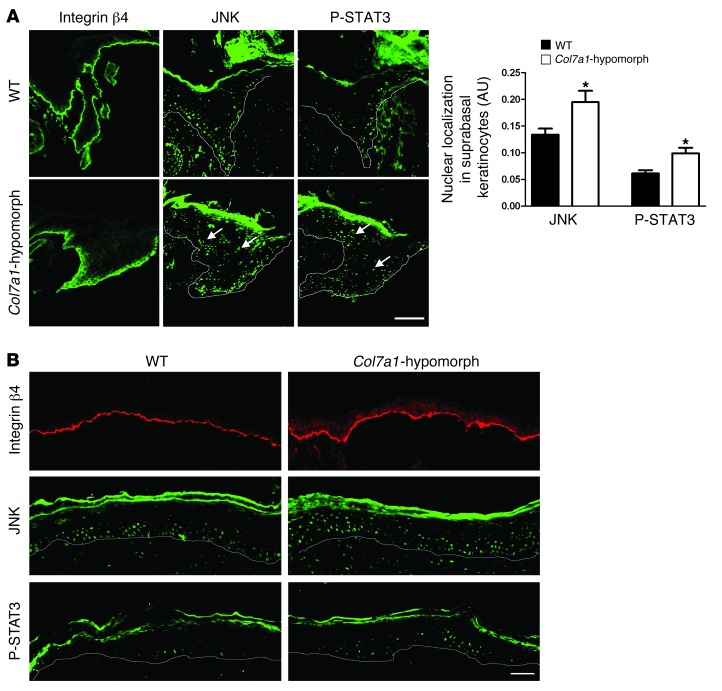
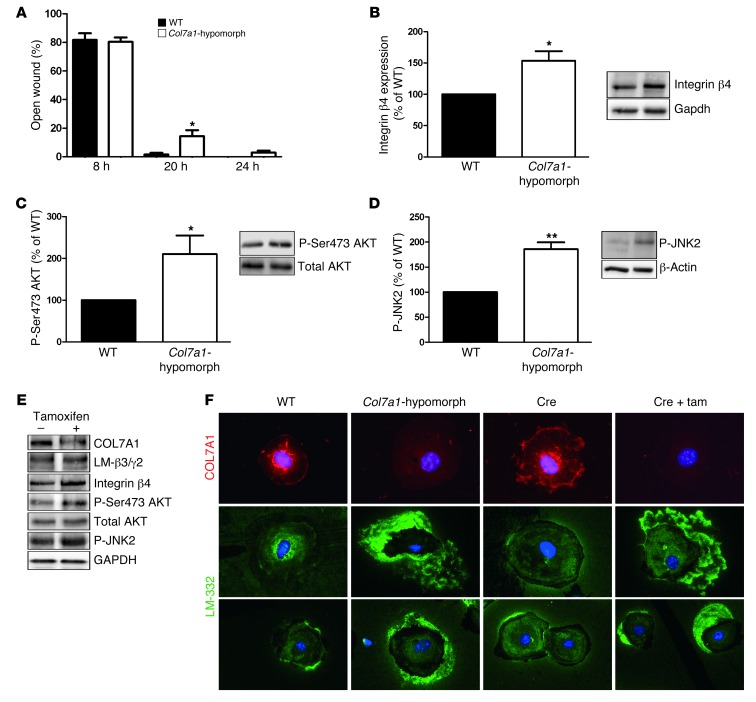
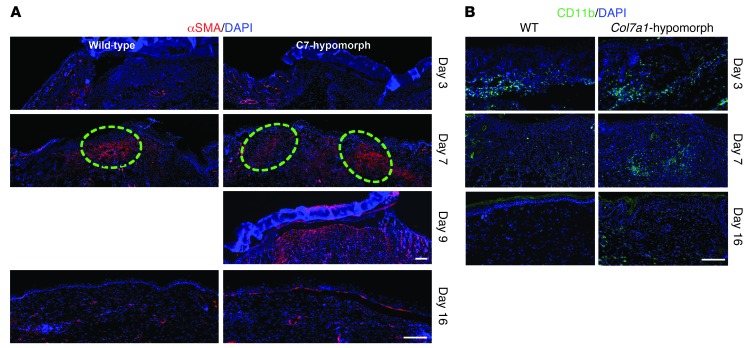
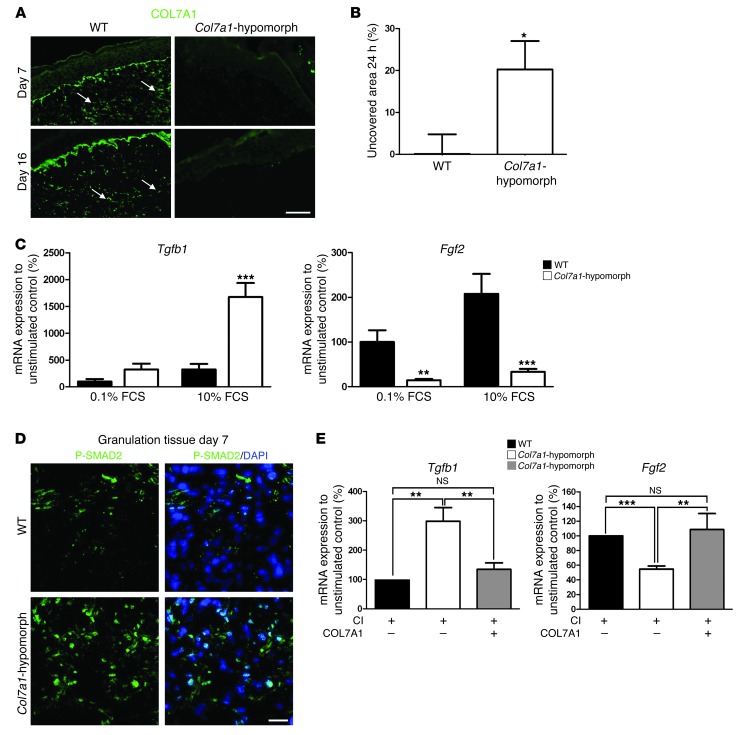
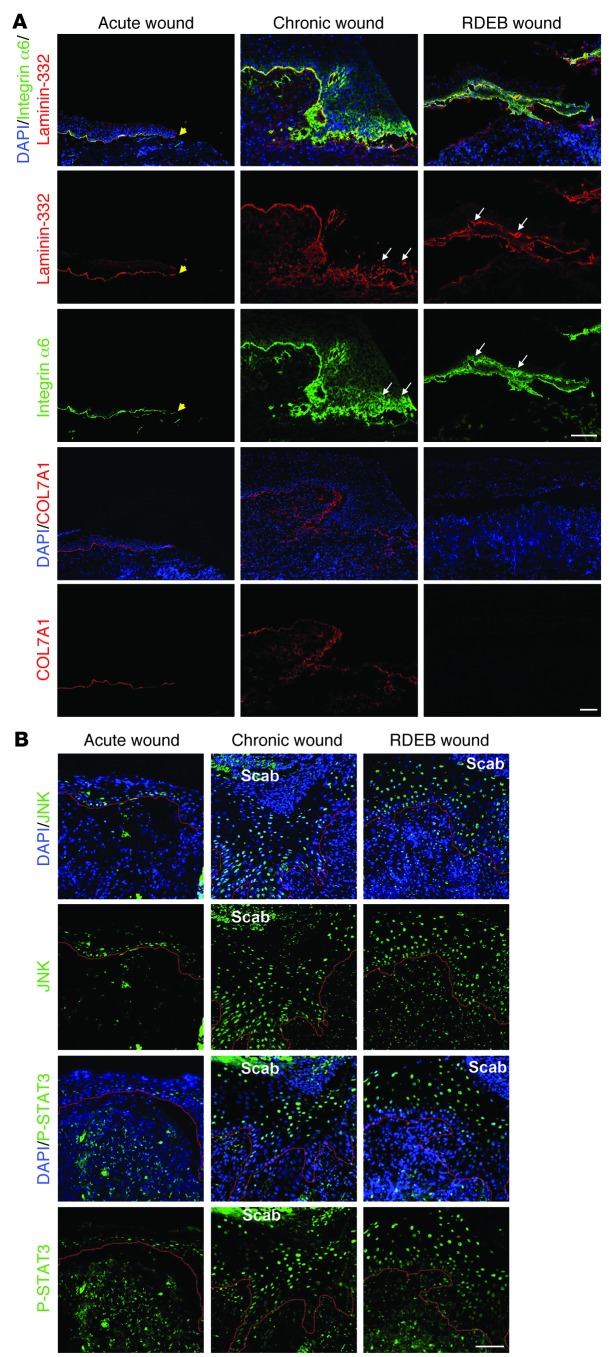
Although a host of intracellular signals is known to contribute to wound healing, the role of the cell microenvironment in tissue repair remains elusive. Here we employed 2 different mouse models of genetic skin fragility to assess the role of the basement membrane protein collagen VII (COL7A1) in wound healing. COL7A1 secures the attachment of the epidermis to the dermis, and its mutations cause a human skin fragility disorder coined recessive dystrophic epidermolysis bullosa (RDEB) that is associated with a constant wound burden. We show that COL7A1 is instrumental for skin wound closure by 2 interconnected mechanisms. First, COL7A1 was required for re-epithelialization through organization of laminin-332 at the dermal-epidermal junction. Its loss perturbs laminin-332 organization during wound healing, which in turn abrogates strictly polarized expression of integrin α6β4 in basal keratinocytes and negatively impacts the laminin-332/integrin α6β4 signaling axis guiding keratinocyte migration. Second, COL7A1 supported dermal fibroblast migration and regulates their cytokine production in the granulation tissue. These findings, which were validated in human wounds, identify COL7A1 as a critical player in physiological wound healing in humans and mice and may facilitate development of therapeutic strategies not only for RDEB, but also for other chronic wounds.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
