

Differentiated roles for MreB-actin isologues and autolytic enzymes in Bacillus subtilis morphogenesis
- PMID: 23869552
- PMCID: PMC3817527
- DOI: 10.1111/mmi.12335
Differentiated roles for MreB-actin isologues and autolytic enzymes in Bacillus subtilis morphogenesis
Abstract
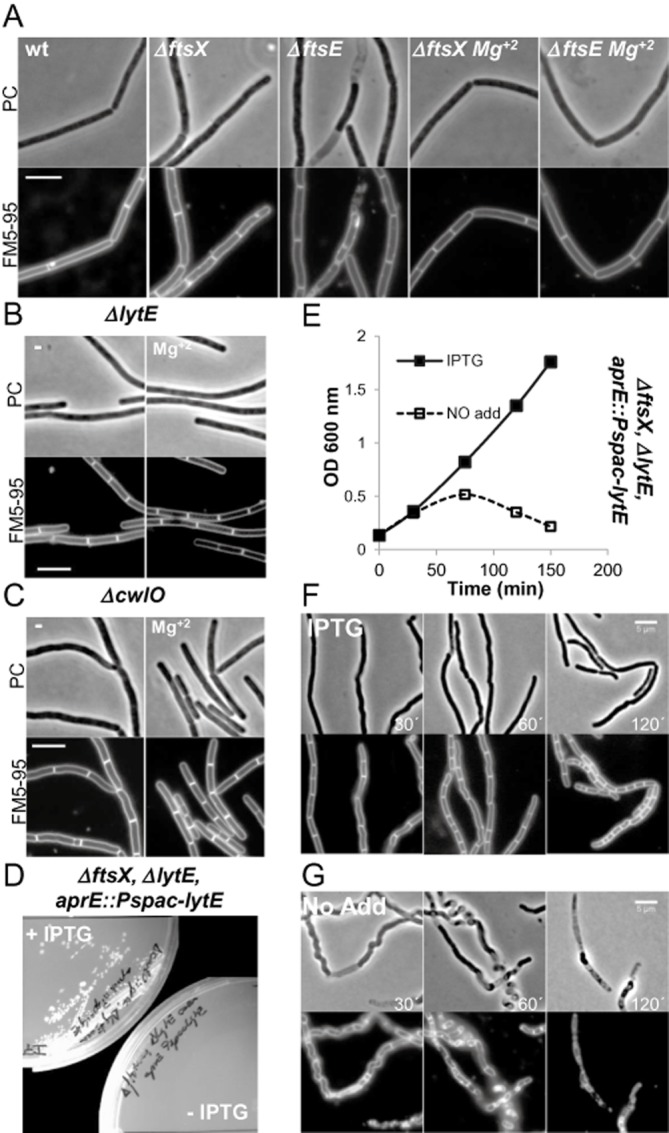
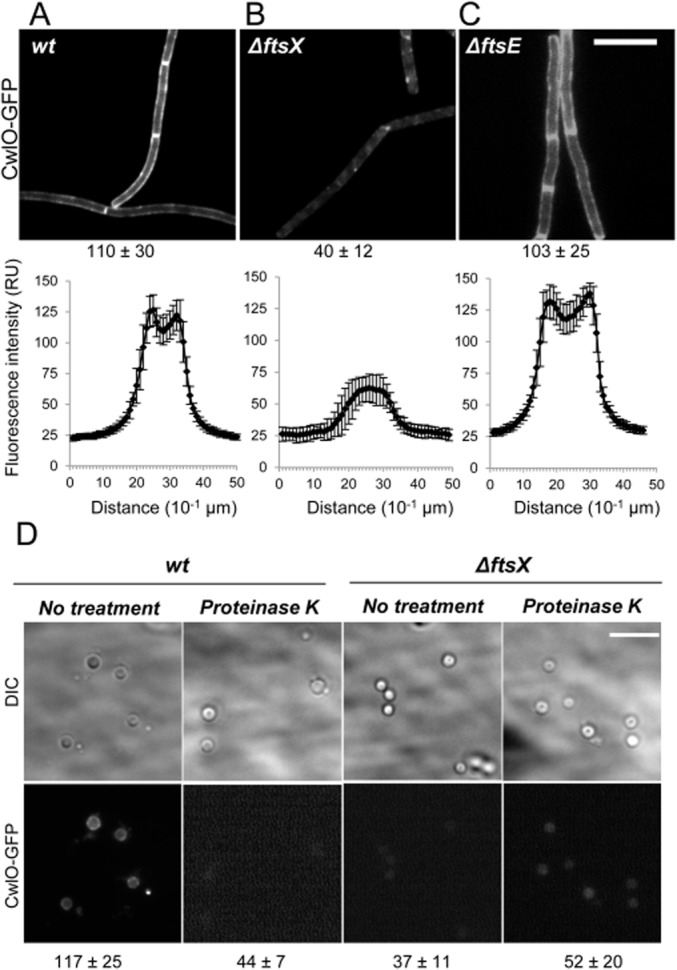
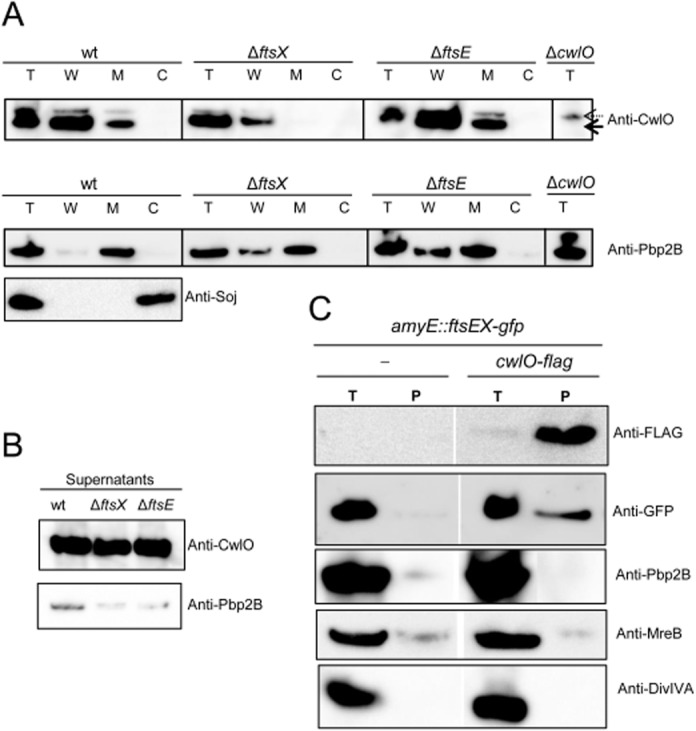
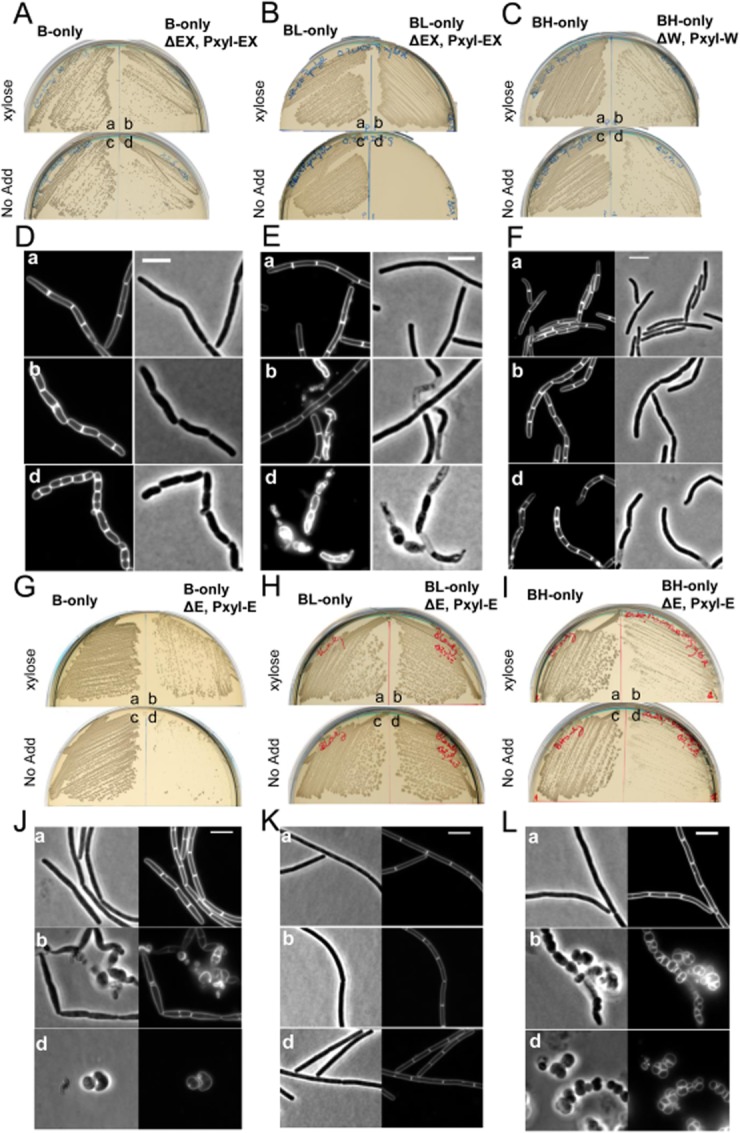
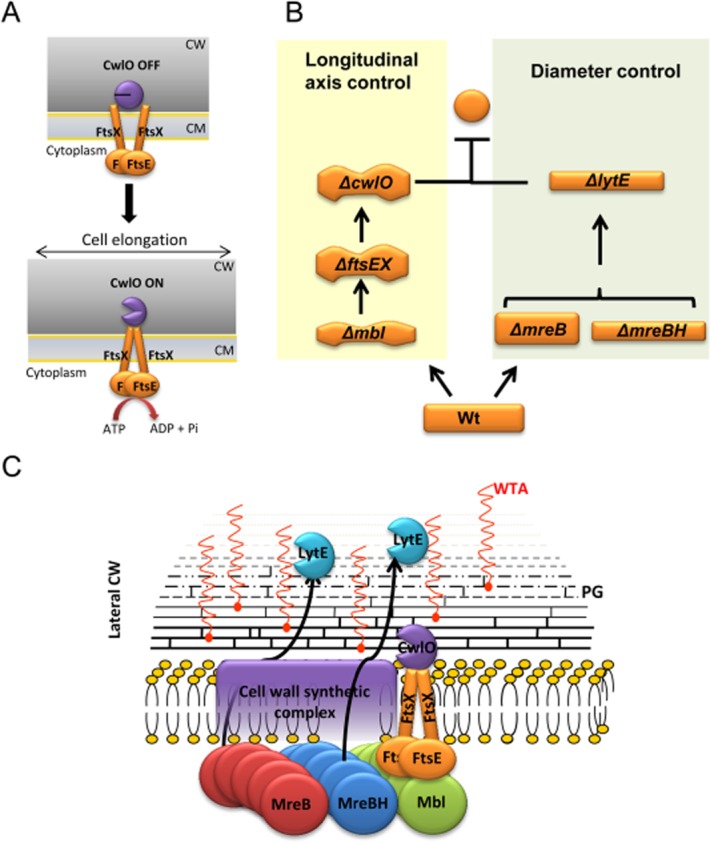
Cell morphogenesis in most bacteria is governed by spatiotemporal growth regulation of the peptidoglycan cell wall layer. Much is known about peptidoglycan synthesis but regulation of its turnover by hydrolytic enzymes is much less well understood. Bacillus subtilis has a multitude of such enzymes. Two of the best characterized are CwlO and LytE: cells lacking both enzymes have a lethal block in cell elongation. Here we show that activity of CwlO is regulated by an ABC transporter, FtsEX, which is required for cell elongation, unlike cell division as in Escherichia coli. Actin-like MreB proteins are thought to play a key role in orchestrating cell wall morphogenesis. B. subtilis has three MreB isologues with partially differentiated functions. We now show that the three MreB isologues have differential roles in regulation of the CwlO and LytE systems and that autolysins control different aspects of cell morphogenesis. The results add major autolytic activities to the growing list of functions controlled by MreB isologues in bacteria and provide new insights into the different specialized functions of essential cell wall autolysins.
© 2013 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Figures

References
-
- Bhavsar AP, Brown ED. Cell wall assembly in Bacillus subtilis: how spirals and spaces challenge paradigms. Mol Microbiol. 2006;60:1077–1090. - PubMed
-
- Bisicchia P, Noone D, Lioliou E, Howell A, Quigley S, Jensen T, et al. The essential YycFG two-component system controls cell wall metabolism in Bacillus subtilis. Mol Microbiol. 2007;65:180–200. - PubMed
-
- den Blaauwen T, de Pedro MA, Nguyen-Disteche M, Ayala JA. Morphogenesis of rod-shaped sacculi. FEMS Microbiol Rev. 2008;32:321–344. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
