

Tandem stem-loops in roX RNAs act together to mediate X chromosome dosage compensation in Drosophila
- PMID: 23870142
- PMCID: PMC3804161
- DOI: 10.1016/j.molcel.2013.07.001
Tandem stem-loops in roX RNAs act together to mediate X chromosome dosage compensation in Drosophila
Abstract
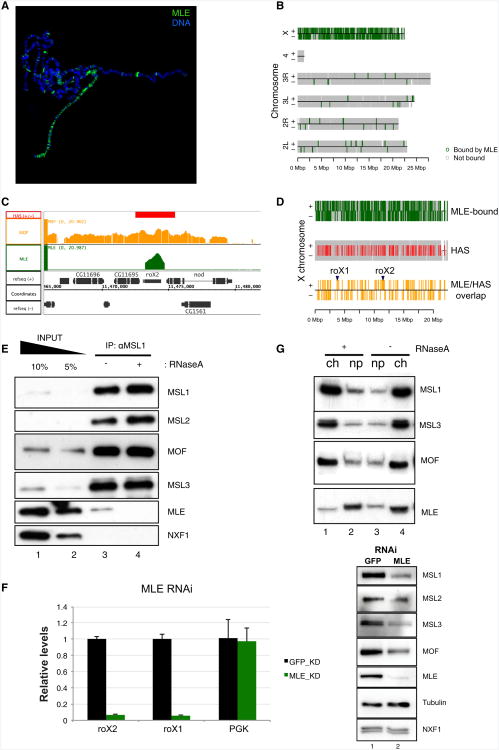
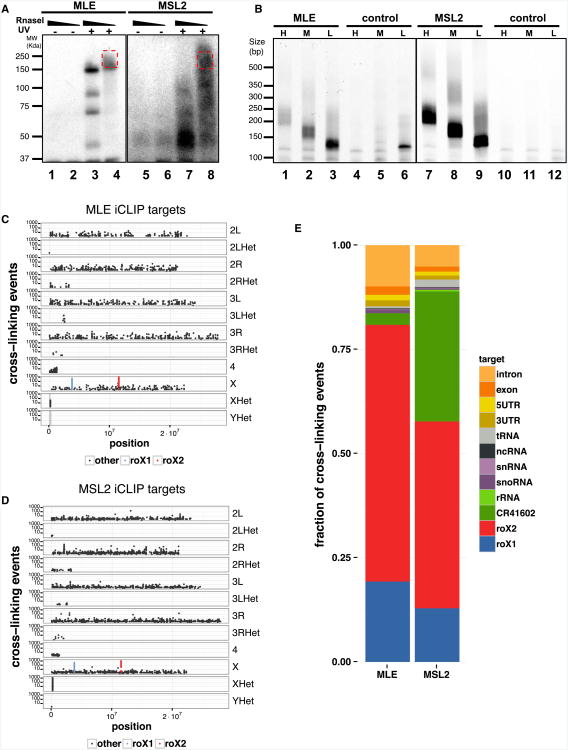
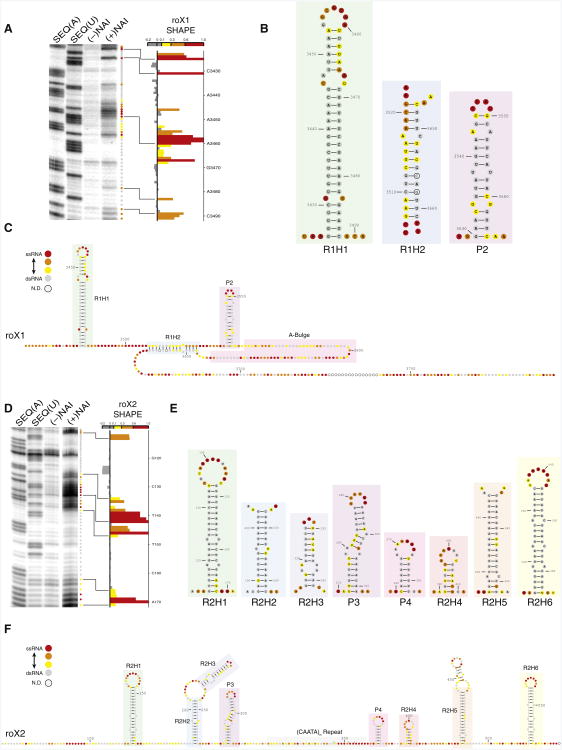
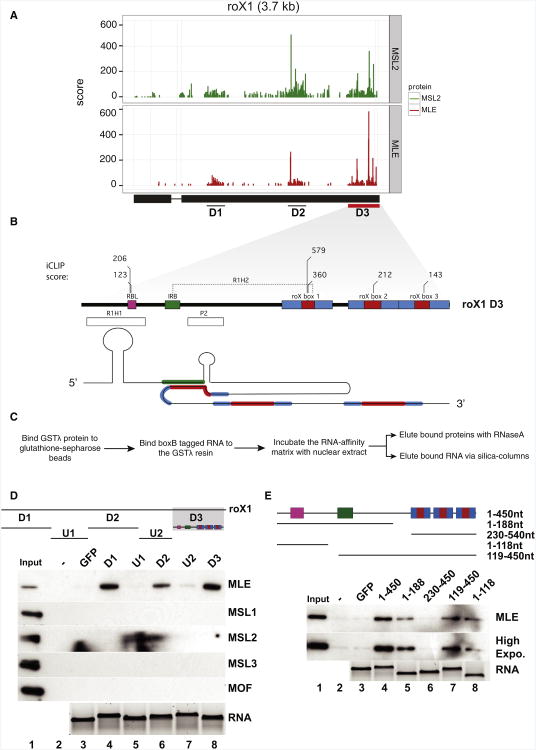
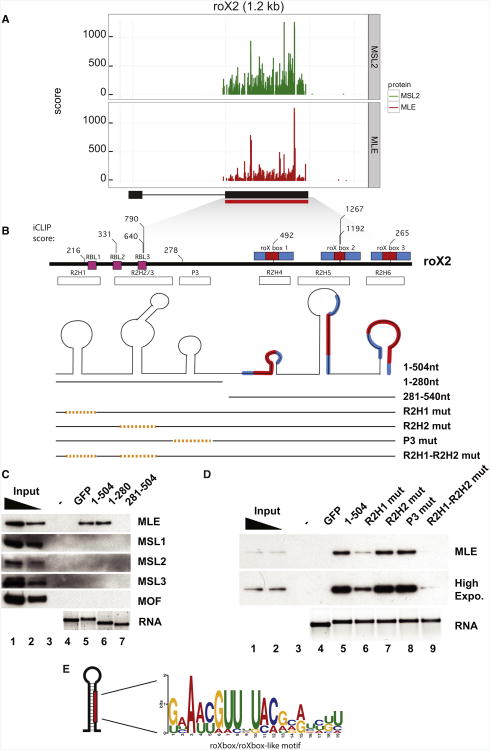
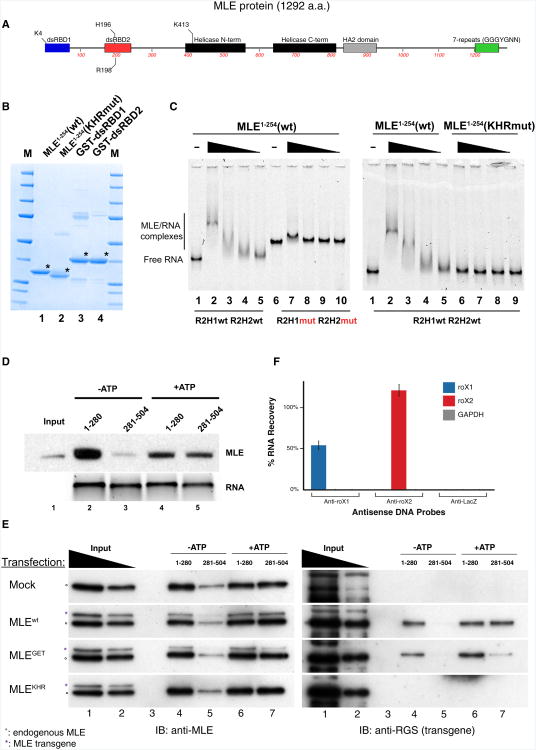
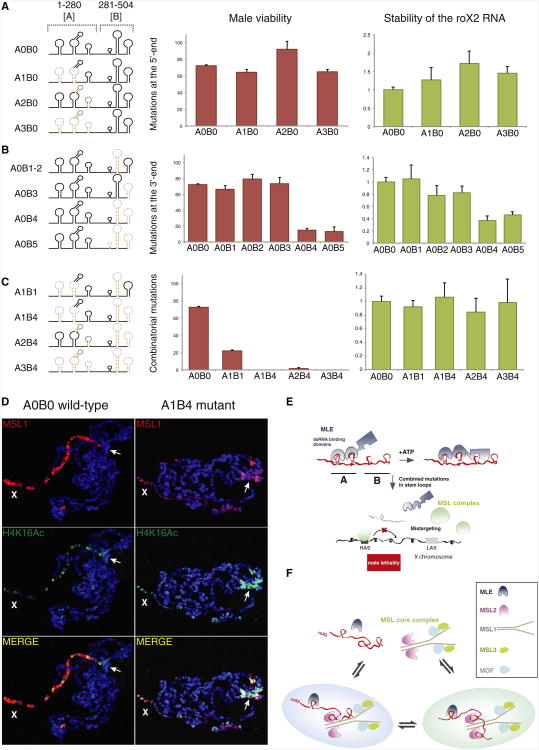
Dosage compensation in Drosophila is an epigenetic phenomenon utilizing proteins and long noncoding RNAs (lncRNAs) for transcriptional upregulation of the male X chromosome. Here, by using UV crosslinking followed by deep sequencing, we show that two enzymes in the Male-Specific Lethal complex, MLE RNA helicase and MSL2 ubiquitin ligase, bind evolutionarily conserved domains containing tandem stem-loops in roX1 and roX2 RNAs in vivo. These domains constitute the minimal RNA unit present in multiple copies in diverse arrangements for nucleation of the MSL complex. MLE binds to these domains with distinct ATP-independent and ATP-dependent behavior. Importantly, we show that different roX RNA domains have overlapping function, since only combinatorial mutations in the tandem stem-loops result in severe loss of dosage compensation and consequently male-specific lethality. We propose that repetitive structural motifs in lncRNAs could provide plasticity during multiprotein complex assemblies to ensure efficient targeting in cis or in trans along chromosomes.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Noncoding roX RNA remodeling triggers fly dosage compensation complex assembly.Mol Cell. 2013 Jul 25;51(2):131-2. doi: 10.1016/j.molcel.2013.07.007. Mol Cell. 2013. PMID: 23870139
-
Non-coding RNA: Structure and function for lncRNAs.Nat Rev Genet. 2013 Sep;14(9):598. doi: 10.1038/nrg3561. Epub 2013 Aug 6. Nat Rev Genet. 2013. PMID: 23917630 No abstract available.
References
-
- Akhtar A, Zink D, Becker PB. Chromodomains are protein-RNA interaction modules. Nature. 2000;407:405–409. - PubMed
-
- Amrein H, Axel R. Genes expressed in neurons of adult male Drosophila. Cell. 1997;88:459–469. - PubMed
-
- Augui S, Nora EP, Heard E. Regulation of X-chromosome inactivation by the X-inactivation centre. Nat Rev Genet. 2011;12:429–442. - PubMed
-
- Conrad T, Akhtar A. Dosage compensation in Drosophila melanogaster: epigenetic fine-tuning of chromosome-wide transcription. Nat Rev Genet. 2011;13:123–134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
