

The DHX33 RNA helicase senses cytosolic RNA and activates the NLRP3 inflammasome
- PMID: 23871209
- PMCID: PMC3756931
- DOI: 10.1016/j.immuni.2013.07.001
The DHX33 RNA helicase senses cytosolic RNA and activates the NLRP3 inflammasome
Abstract
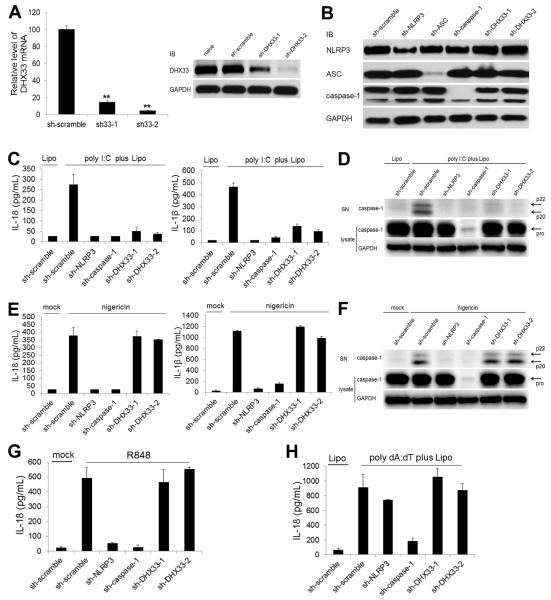
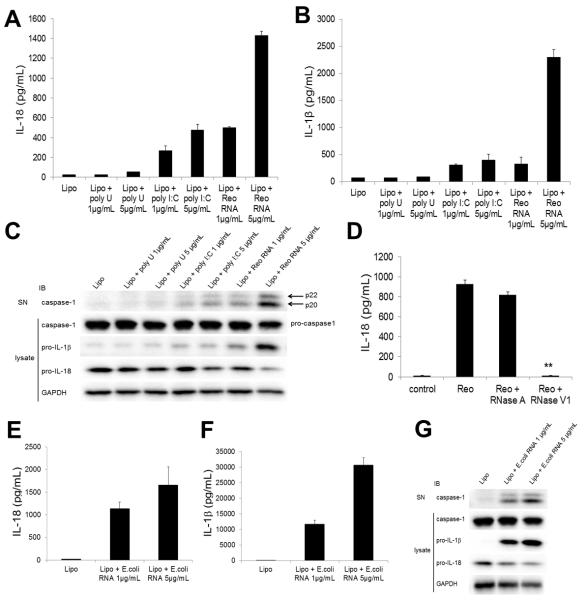
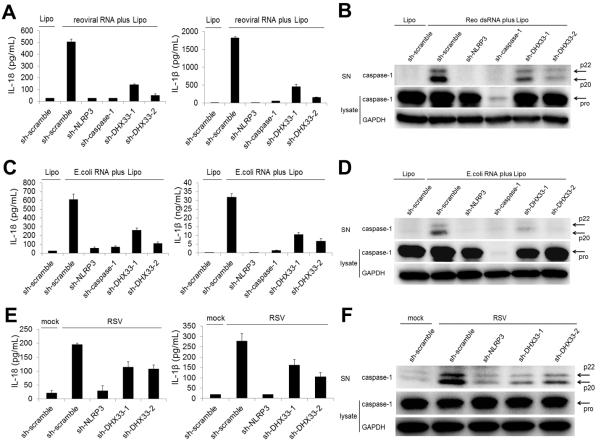
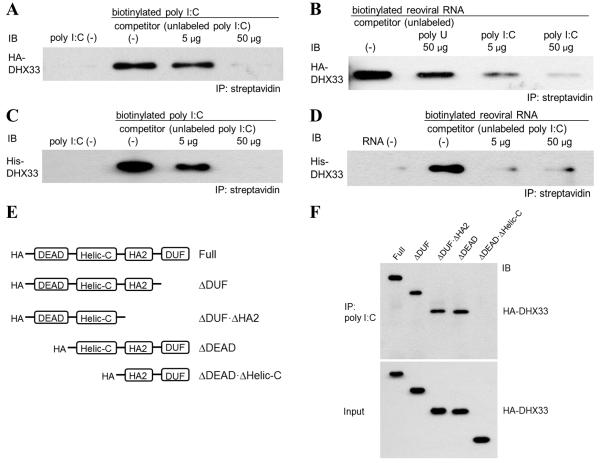
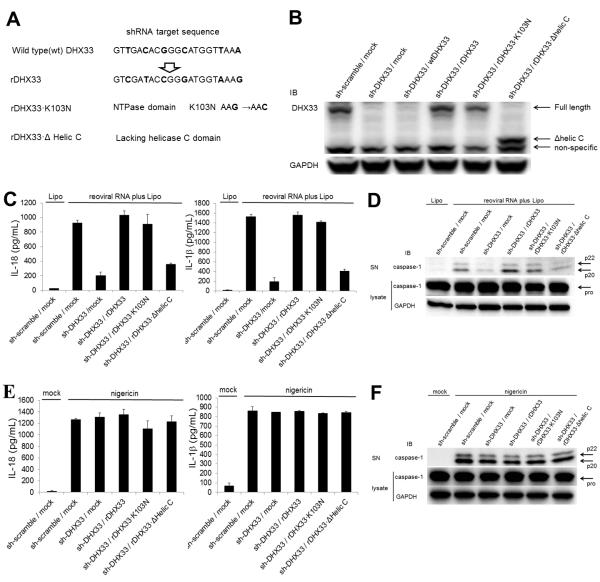
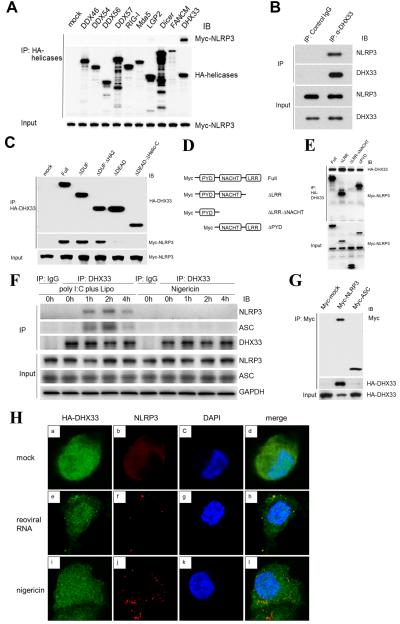
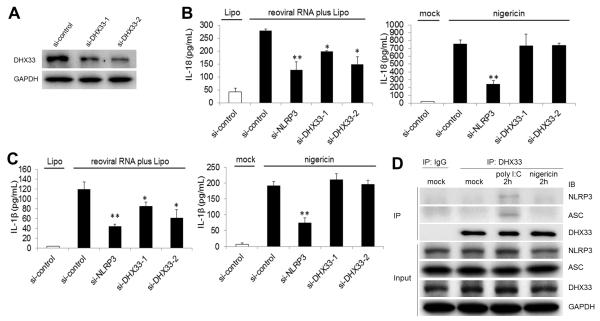
The NLRP3 inflammasome plays a major role in innate immune responses by activating caspase-1, resulting in secretion of interleukin-18 (IL-18) and IL-1β. Although cytosolic double-stranded RNA (dsRNA) and bacterial RNA are known to activate the NLRP3 inflammasome, the upstream sensor is unknown. We investigated the potential function of DExD/H-box RNA helicase family members (previously shown to sense cytosolic DNA and RNA to induce type 1 interferon responses) in RNA-induced NLRP3 inflammasome activation. Among the helicase family members tested, we found that targeting of DHX33 expression by short hairpin RNA efficiently blocked the activation of caspase-1 and secretion of IL-18 and IL-1β in human macrophages that were activated by cytosolic poly I:C, reoviral RNA, or bacterial RNA. DHX33 bound dsRNA via the helicase C domain. DHX33 interacted with NLRP3 and formed the inflammasome complex following stimulation with RNA. We therefore identified DHX33 as a cytosolic RNA sensor that activates the NLRP3 inflammasome.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
RNA helicase DHX33 puts a new twist on NLRP3 inflammasome activation.Immunity. 2013 Jul 25;39(1):94-6. doi: 10.1016/j.immuni.2013.07.010. Immunity. 2013. PMID: 23890068
References
-
- Besch R, Poeck H, Hohenauer T, Senft D, Hacker G, Berking C, Hornung V, Endres S, Ruzicka T, Rothenfusser S, Hartmann G. Proapoptotic signaling induced by RIG-I and MDA-5 results in type I interferon-independent apoptosis in human melanoma cells. The Journal of clinical investigation. 2009;119:2399–2411. - PMC - PubMed
-
- Bryant C, Fitzgerald KA. Molecular mechanisms involved in inflammasome activation. Trends in cell biology. 2009;19:455–464. - PubMed
-
- Burckstummer T, Baumann C, Bluml S, Dixit E, Durnberger G, Jahn H, Planyavsky M, Bilban M, Colinge J, Bennett KL, Superti-Furga G. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nature immunology. 2009;10:266–272. - PubMed
-
- Eigenbrod T, Franchi L, Munoz-Planillo R, Kirschning CJ, Freudenberg MA, Nunez G, Dalpke A. Bacterial RNA mediates activation of caspase-1 and IL-1beta release independently of TLRs 3, 7, 9 and TRIF but is dependent on UNC93B. Journal of immunology (Baltimore, Md.: 1950) 2012;189:328–336. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
