

TMC1 and TMC2 are components of the mechanotransduction channel in hair cells of the mammalian inner ear
- PMID: 23871232
- PMCID: PMC3827726
- DOI: 10.1016/j.neuron.2013.06.019
TMC1 and TMC2 are components of the mechanotransduction channel in hair cells of the mammalian inner ear
Abstract
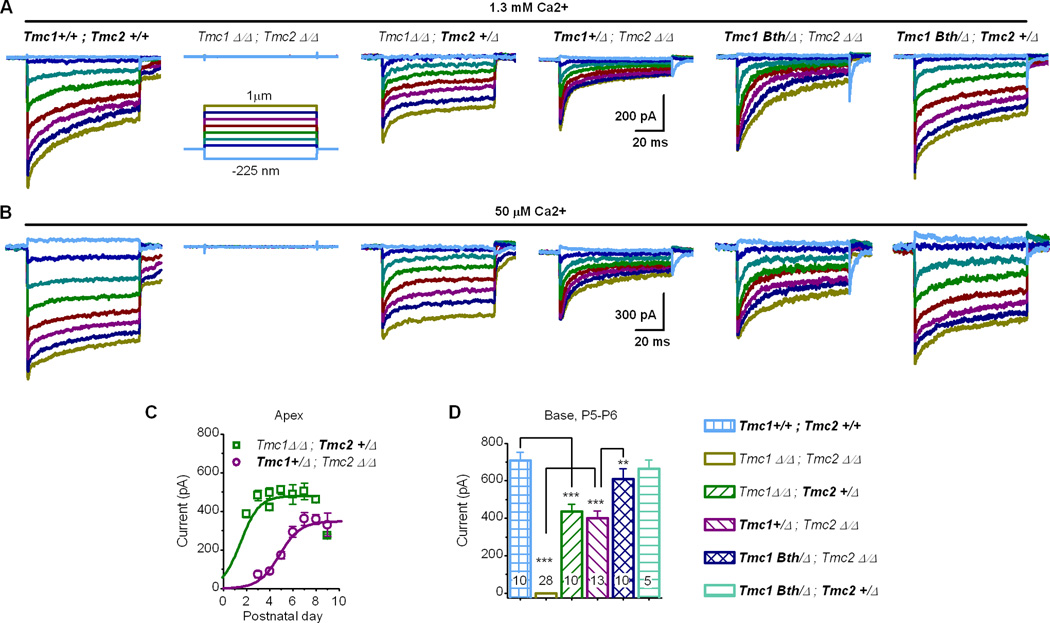
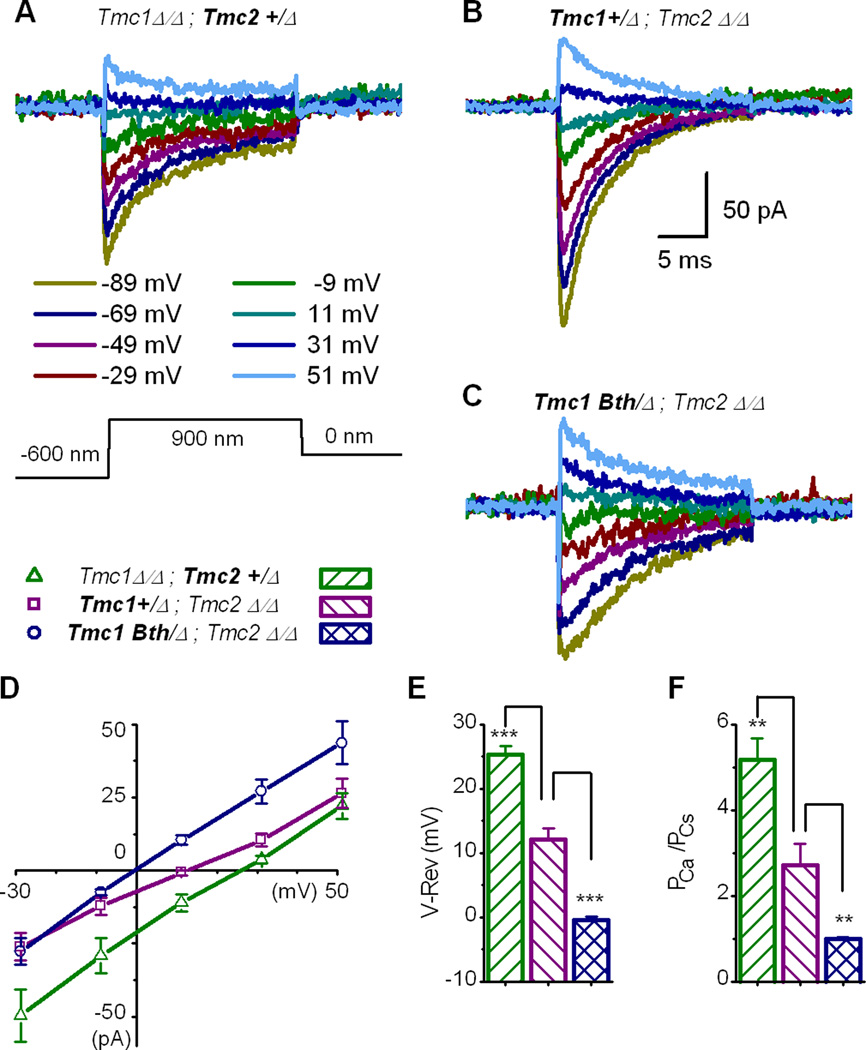
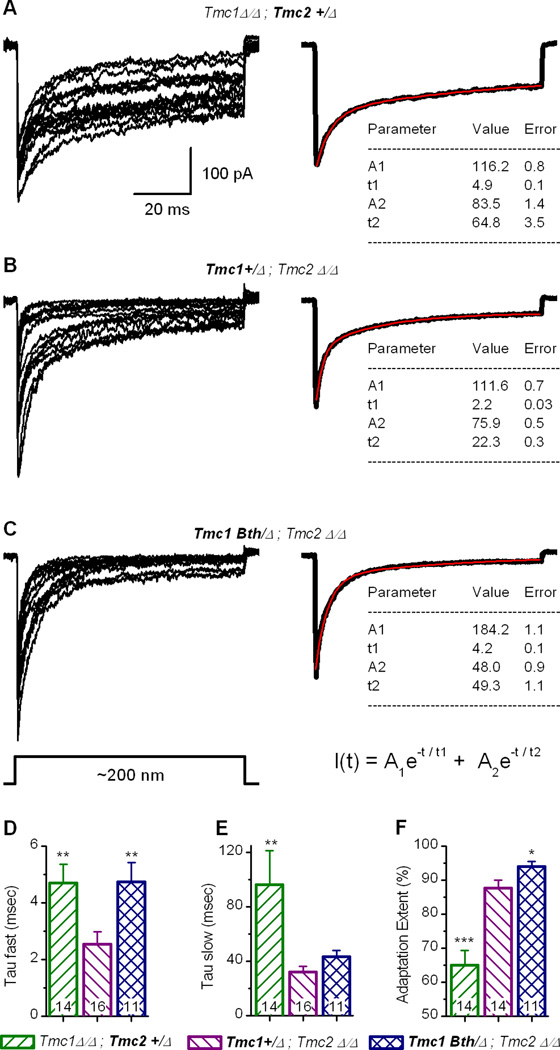
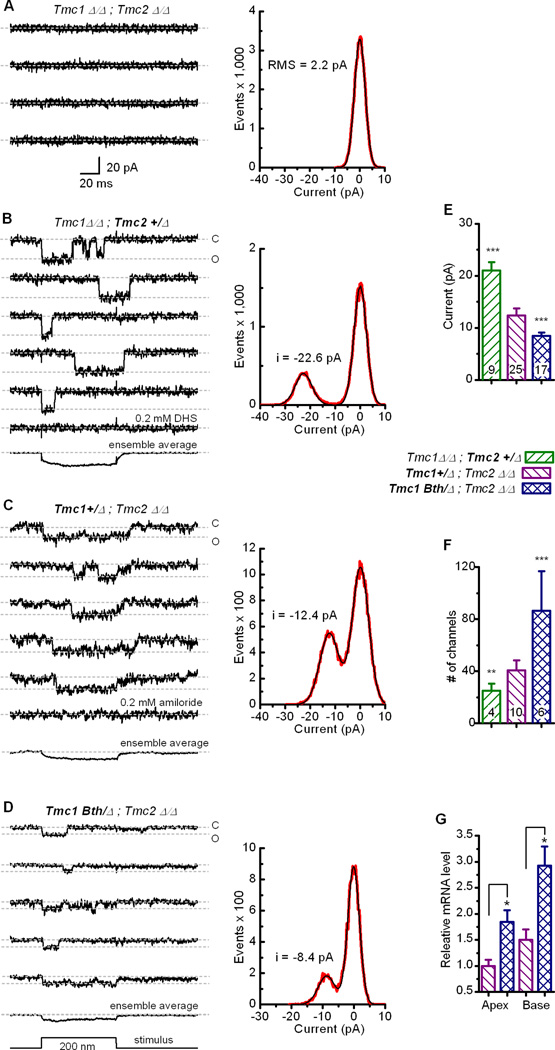
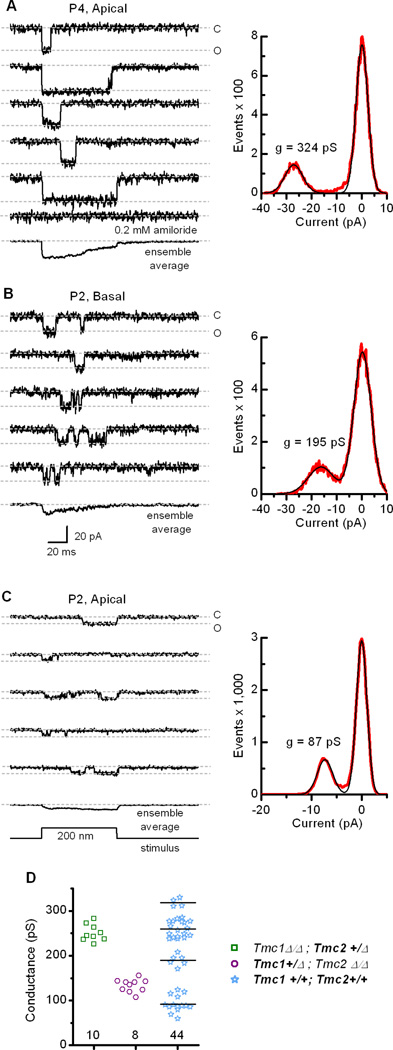
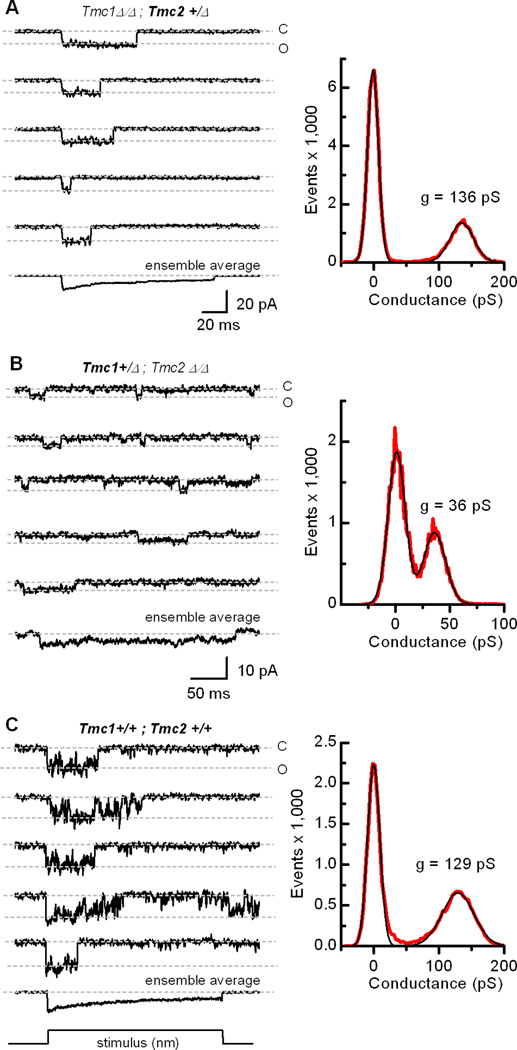
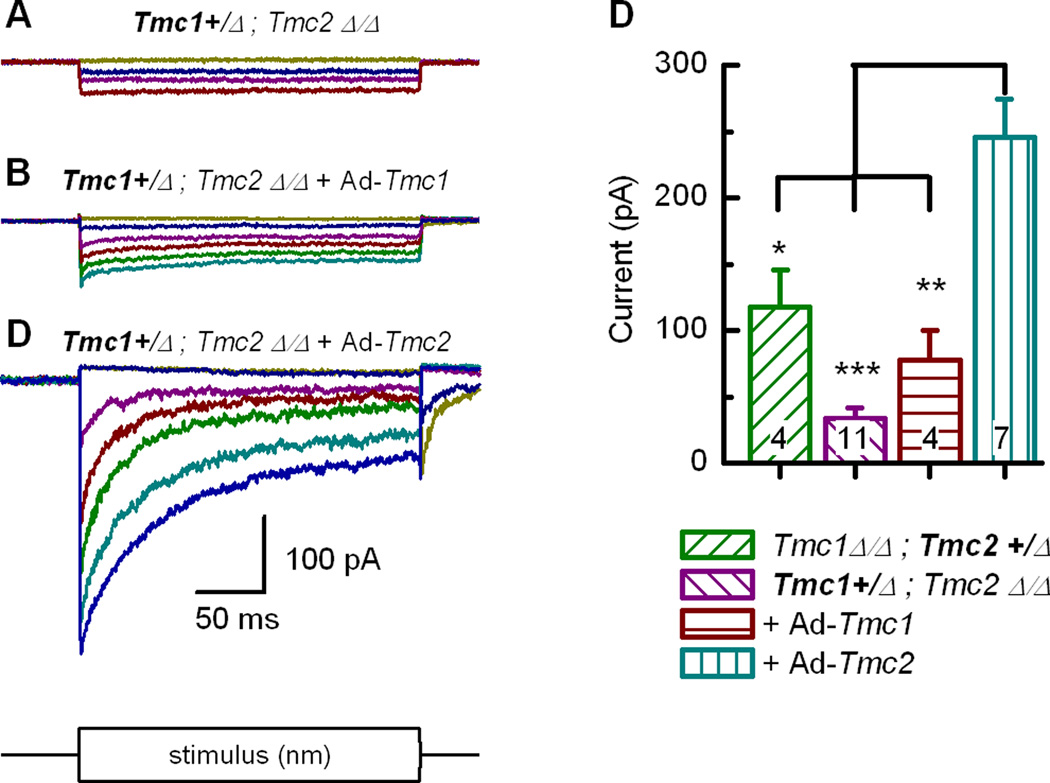
Sensory transduction in auditory and vestibular hair cells requires expression of transmembrane channel-like (Tmc) 1 and 2 genes, but the function of these genes is unknown. To investigate the hypothesis that TMC1 and TMC2 proteins are components of the mechanosensitive ion channels that convert mechanical information into electrical signals, we recorded whole-cell and single-channel currents from mouse hair cells that expressed Tmc1, Tmc2, or mutant Tmc1. Cells that expressed Tmc2 had high calcium permeability and large single-channel currents, while cells with mutant Tmc1 had reduced calcium permeability and reduced single-channel currents. Cells that expressed Tmc1 and Tmc2 had a broad range of single-channel currents, suggesting multiple heteromeric assemblies of TMC subunits. The data demonstrate TMC1 and TMC2 are components of hair cell transduction channels and contribute to permeation properties. Gradients in TMC channel composition may also contribute to variation in sensory transduction along the tonotopic axis of the mammalian cochlea.
Copyright © 2013 Elsevier Inc. All rights reserved.
Conflict of interest statement
We disclose the following potential conflicts: Drs. Kurima and Griffith hold U.S. Patents: 7,166,433 (Transductin-2 and Applications to Hereditary Deafness; abandoned in 2009), 7,192,705 (Transductin-1 and Applications to Hereditary Deafness) and 7,659,115 (Nucleic Acid Encoding Human Transductin-1 Polypeptide). These patents have never been licensed and generate no income or royalties. There are no other conflicts.
Figures
Comment in
-
Mechanotransduction: the elusive hair cell transduction channel revealed?Curr Biol. 2013 Oct 7;23(19):R887-90. doi: 10.1016/j.cub.2013.08.031. Curr Biol. 2013. PMID: 24112987
References
-
- Arnadóttir J, Chalfie M. Eukaryotic mechanosensitive channels. Annu. Rev. Biophys. 2010;39:111–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
