

Cell-to-cell heterogeneity in lipid droplets suggests a mechanism to reduce lipotoxicity
- PMID: 23871243
- PMCID: PMC3746173
- DOI: 10.1016/j.cub.2013.06.032
Cell-to-cell heterogeneity in lipid droplets suggests a mechanism to reduce lipotoxicity
Abstract
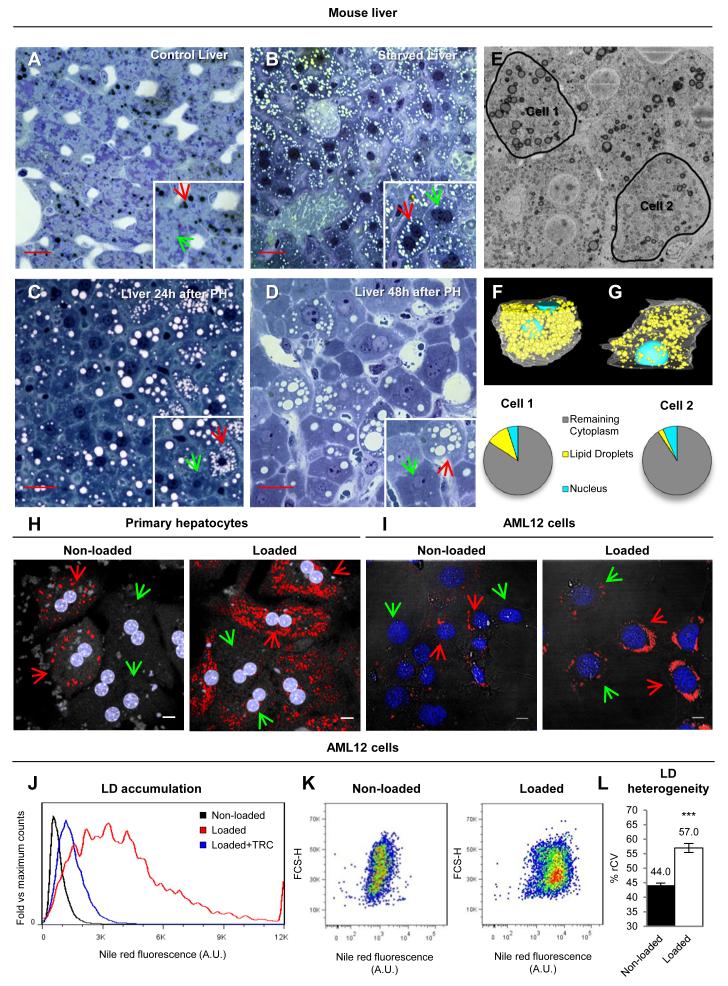
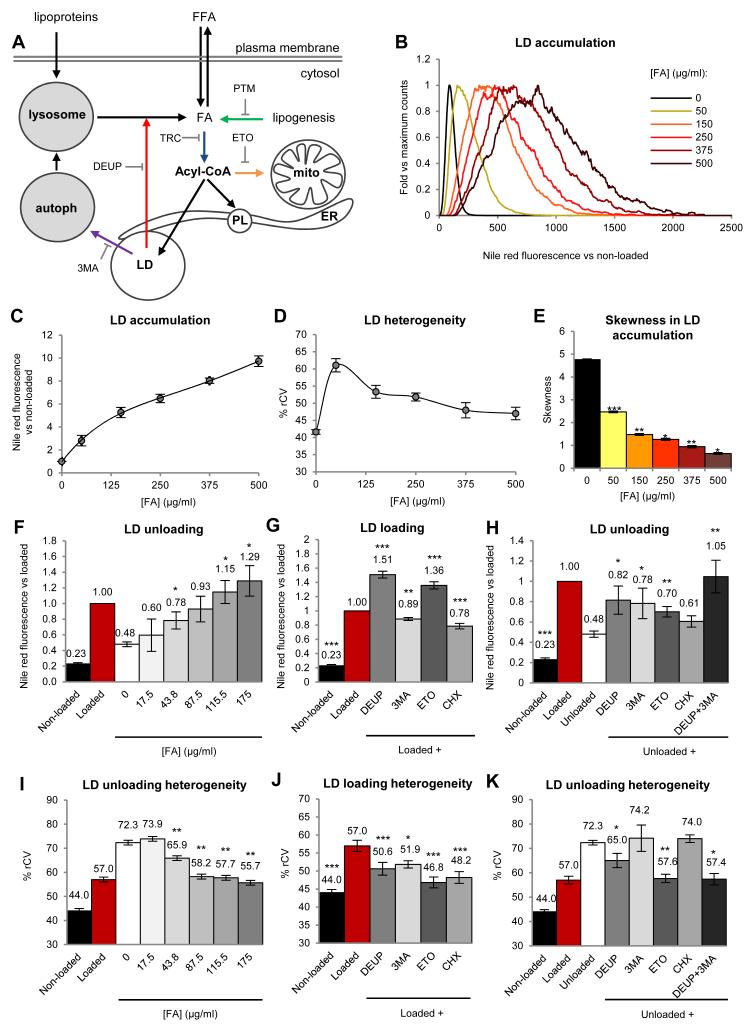
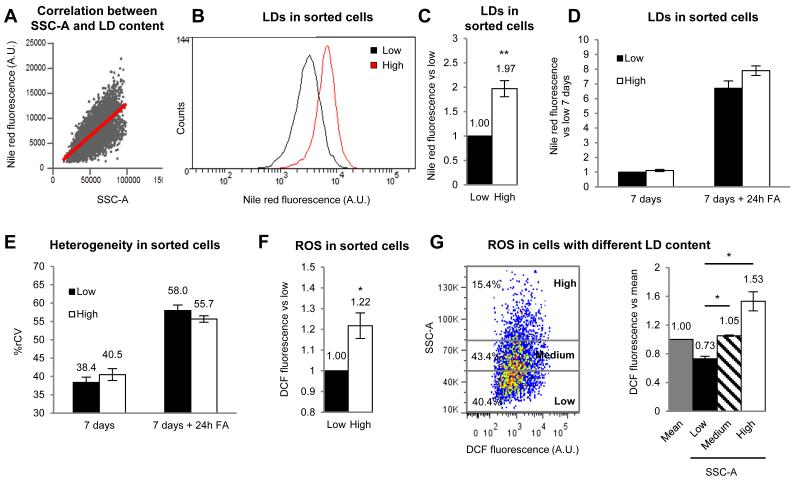
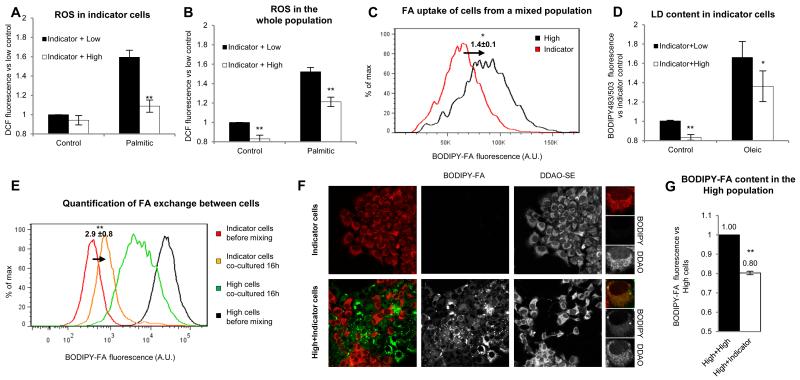
Lipid droplets (LDs) are dynamic organelles that collect, store, and supply lipids [1]. LDs have a central role in the exchange of lipids occurring between the cell and the environment and provide cells with substrates for energy metabolism, membrane synthesis, and production of lipid-derived molecules such as lipoproteins or hormones. However, lipid-derived metabolites also cause progressive lipotoxicity [2], accumulation of reactive oxygen species (ROS), endoplasmic reticulum stress, mitochondrial malfunctioning, and cell death [2]. Intracellular accumulation of LDs is a hallmark of prevalent human diseases, including obesity, steatosis, diabetes, myopathies, and arteriosclerosis [3]. Indeed, nonalcoholic fatty liver disease is the most common cause of abnormal hepatic function among adults [4, 5]. Lipotoxicity gradually promotes cellular ballooning and disarray, megamitochondria, accumulation of Mallory's hyaline in hepatocytes, and inflammation, fibrosis, and cirrhosis in the liver. Here, using confocal microscopy, serial-block-face scanning electron microscopy, and flow cytometry, we show that LD accumulation is heterogeneous within a cell population and follows a positive skewed distribution. Lipid availability and fluctuations in biochemical networks controlling lipolysis, fatty acid oxidation, and protein synthesis contribute to cell-to-cell heterogeneity. Critically, this reversible variability generates a subpopulation of cells that effectively collect and store lipids. This high-lipid subpopulation accumulates more LDs and more ROS and reduces the risk of lipotoxicity to the population without impairing overall lipid homeostasis, since high-lipid cells can supply stored lipids to the other cells. In conclusion, we demonstrate fat storage compartmentalization within a cell population and propose that this is a protective social organization to reduce lipotoxicity.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Fat accretion in a subpopulation of hepatocytes as a strategy to protect the whole liver against oxidative stress and lipotoxicity.Clin Res Hepatol Gastroenterol. 2013 Dec;37(6):553-5. doi: 10.1016/j.clinre.2013.08.005. Epub 2013 Sep 25. Clin Res Hepatol Gastroenterol. 2013. PMID: 24075194
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
