

NSun2-mediated cytosine-5 methylation of vault noncoding RNA determines its processing into regulatory small RNAs
- PMID: 23871666
- PMCID: PMC3730056
- DOI: 10.1016/j.celrep.2013.06.029
NSun2-mediated cytosine-5 methylation of vault noncoding RNA determines its processing into regulatory small RNAs
Abstract
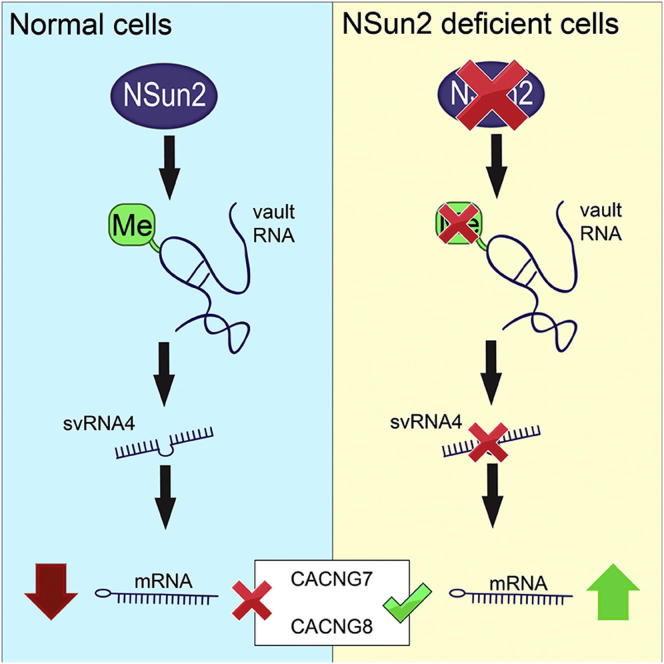
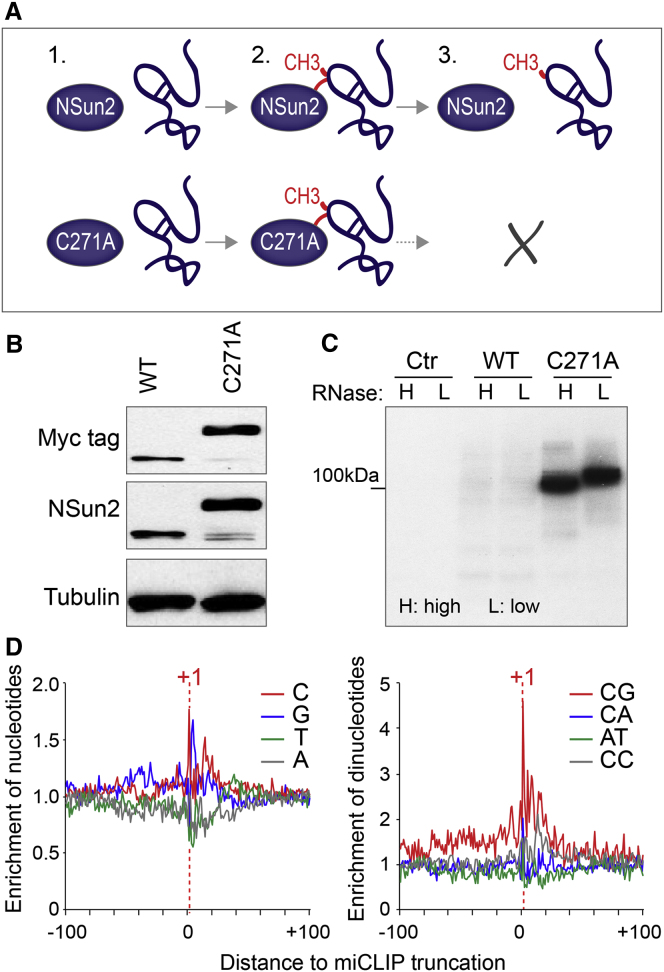
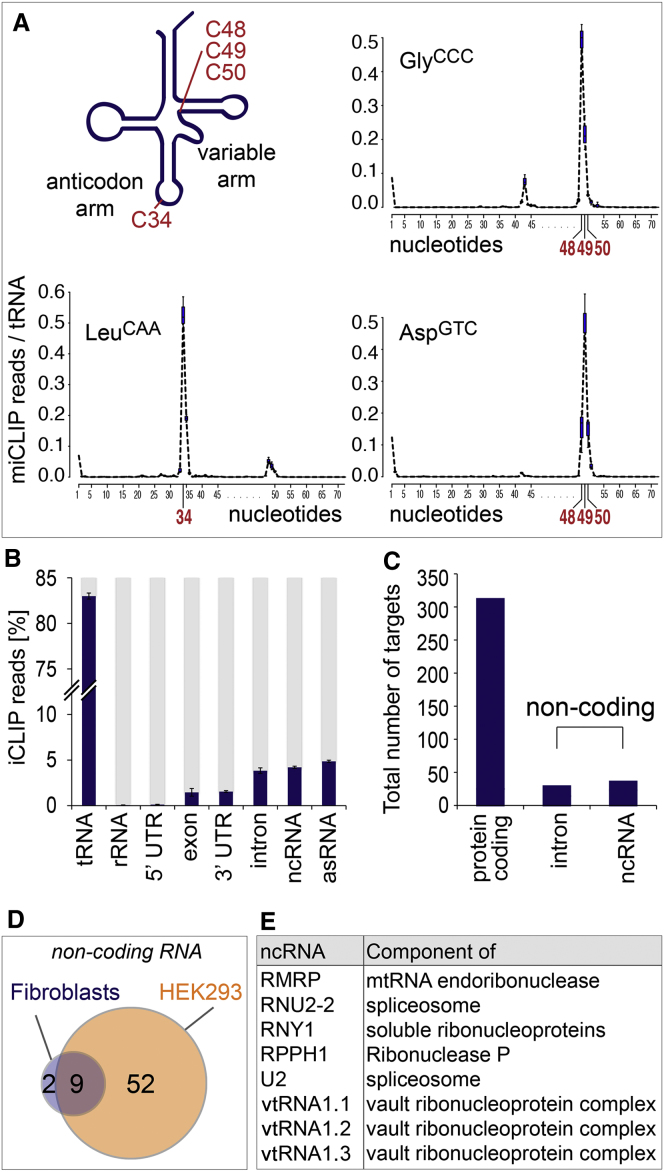
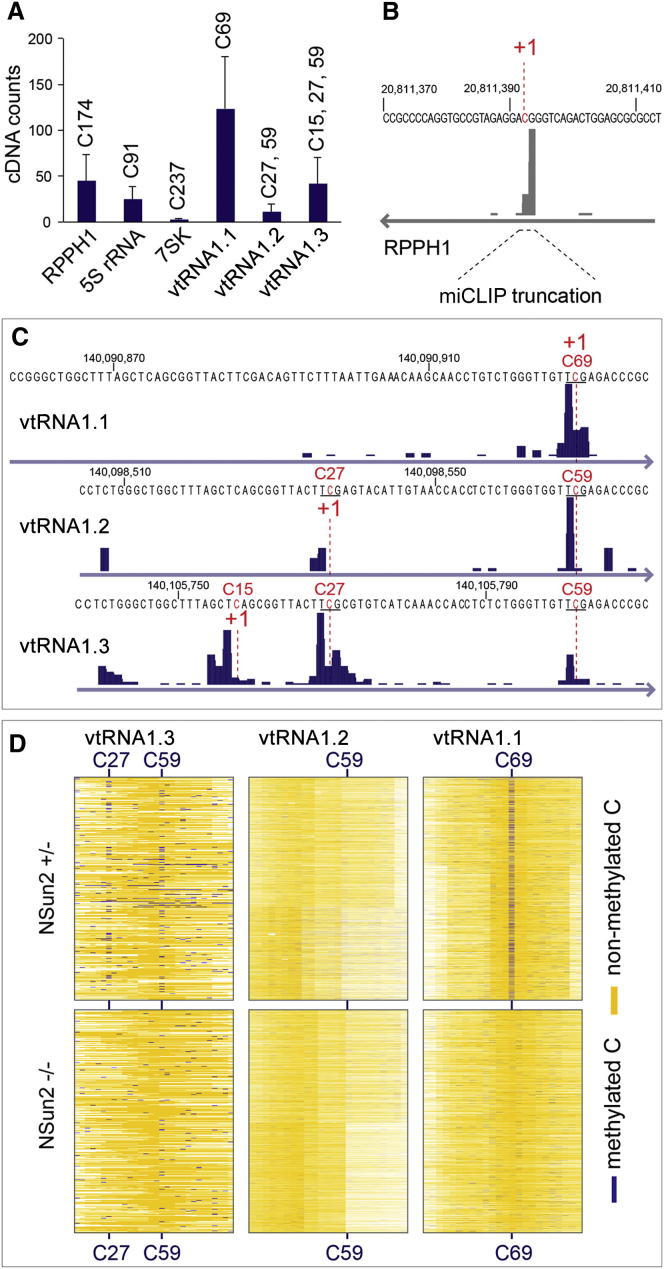
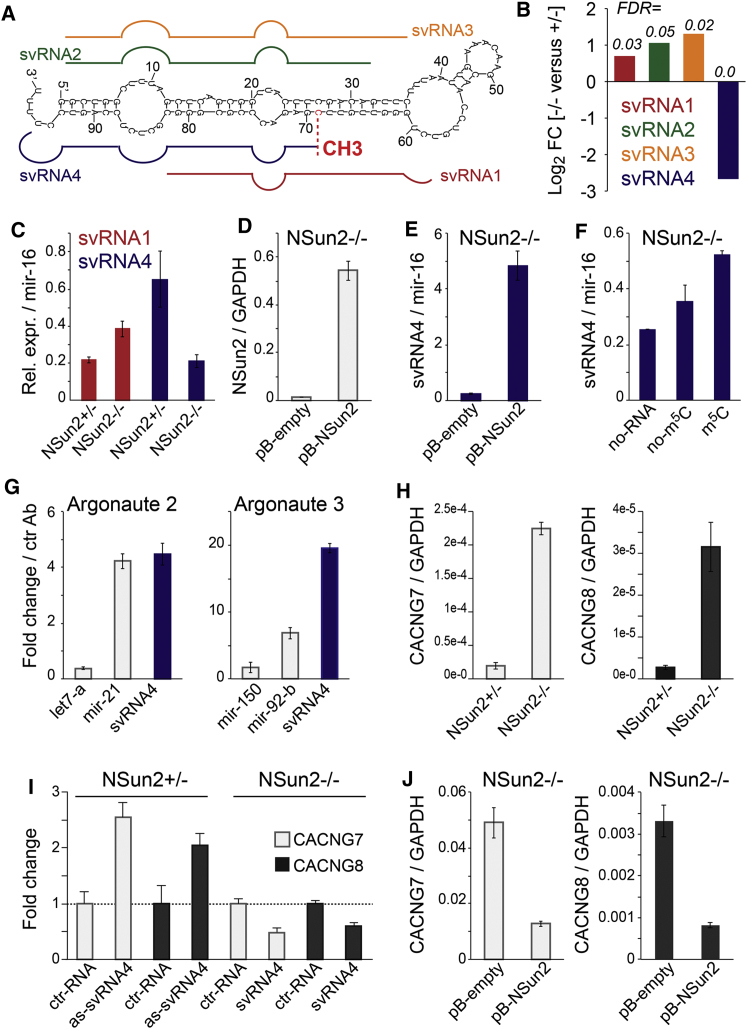
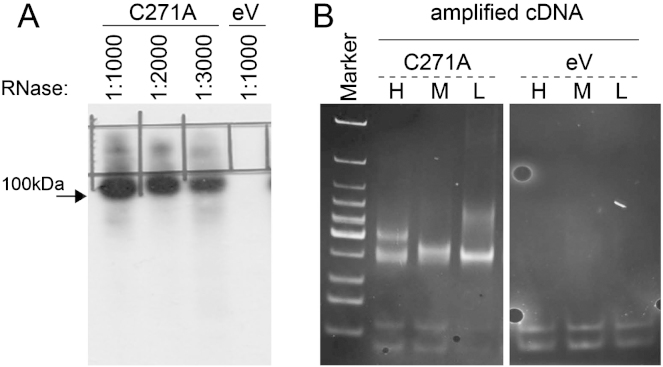
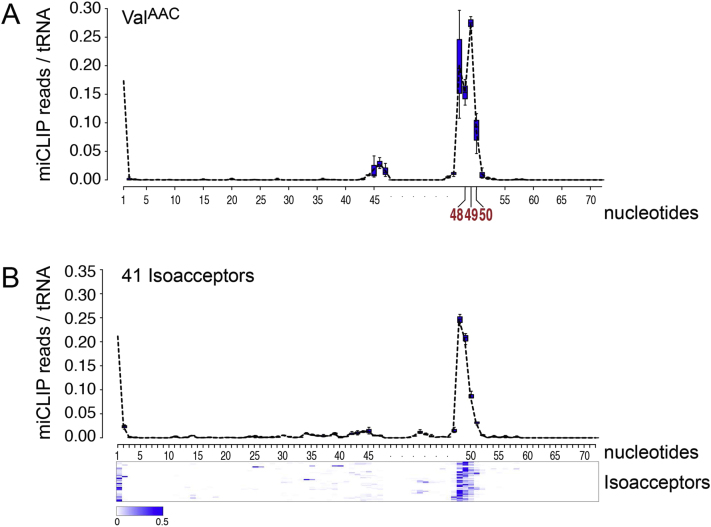
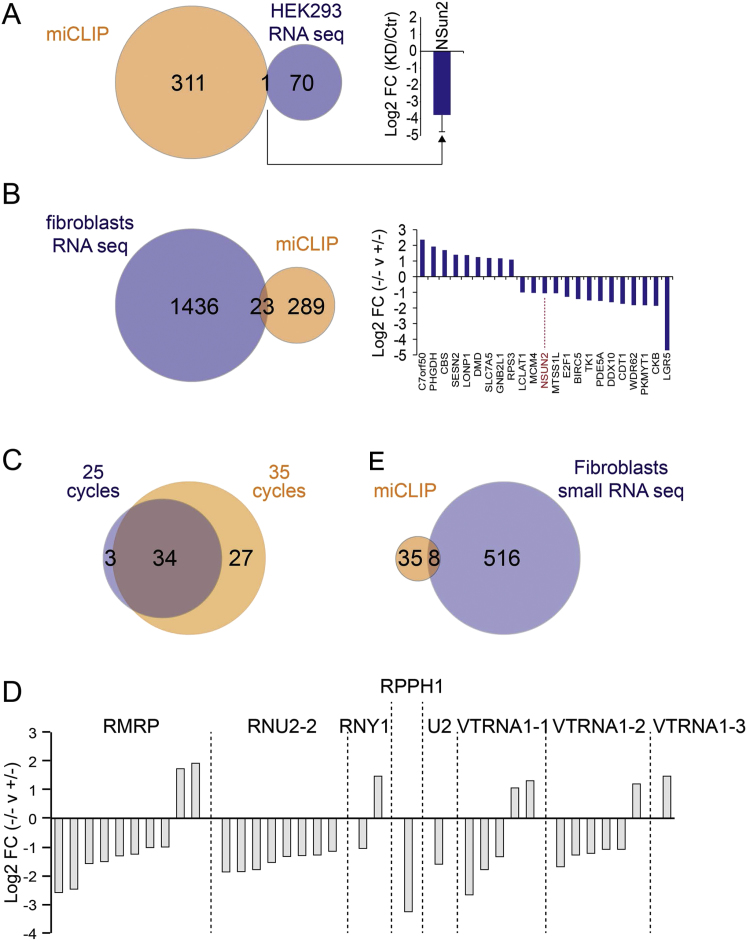
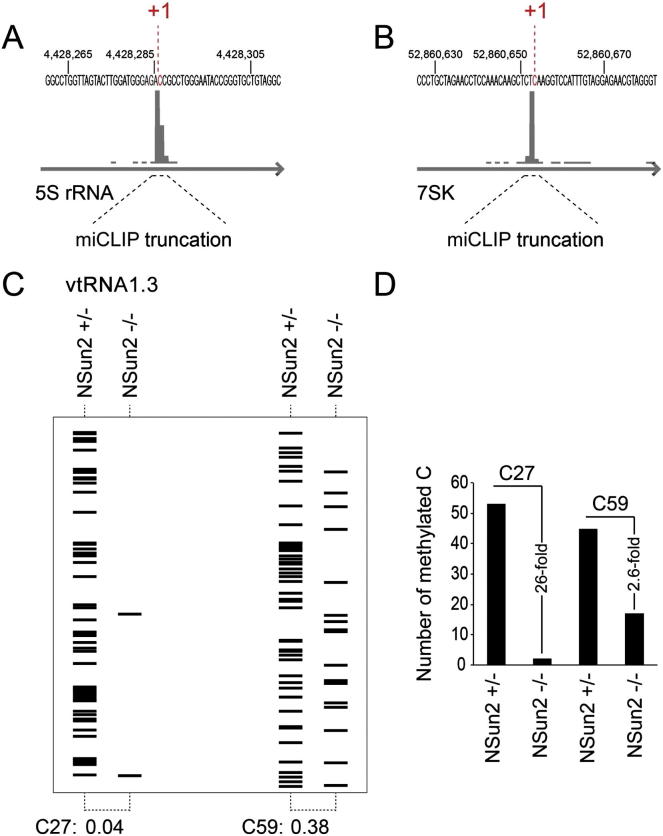
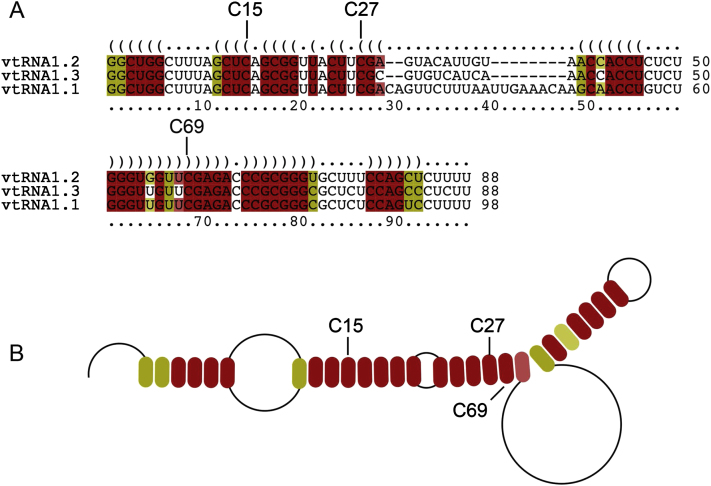
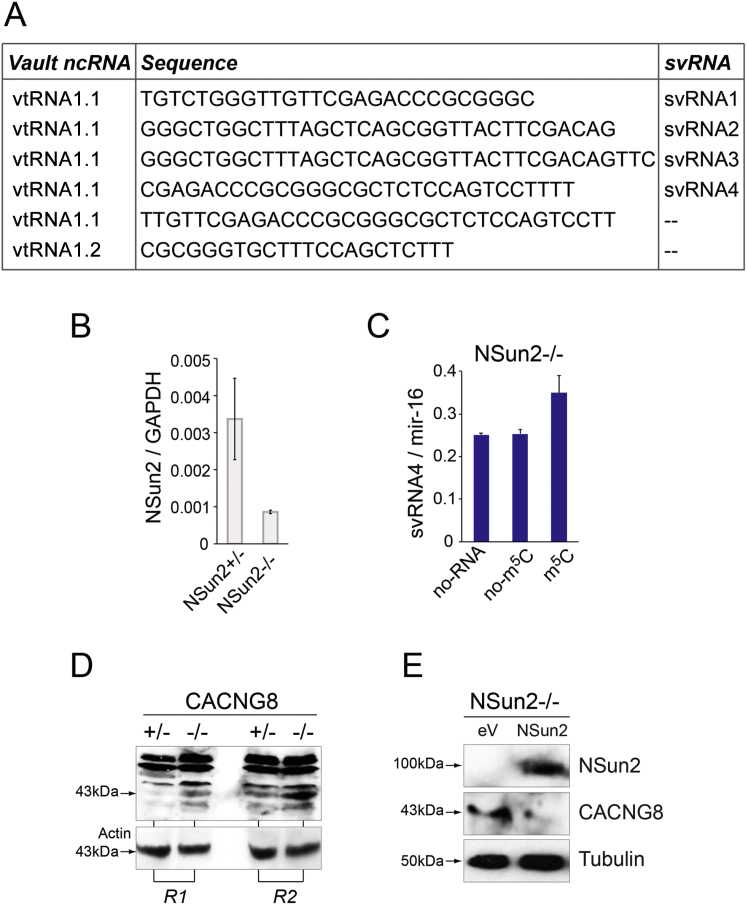
Autosomal-recessive loss of the NSUN2 gene has been identified as a causative link to intellectual disability disorders in humans. NSun2 is an RNA methyltransferase modifying cytosine-5 in transfer RNAs (tRNAs), yet the identification of cytosine methylation in other RNA species has been hampered by the lack of sensitive and reliable molecular techniques. Here, we describe miCLIP as an additional approach for identifying RNA methylation sites in transcriptomes. miCLIP is a customized version of the individual-nucleotide-resolution crosslinking and immunoprecipitation (iCLIP) method. We confirm site-specific methylation in tRNAs and additional messenger and noncoding RNAs (ncRNAs). Among these, vault ncRNAs contained six NSun2-methylated cytosines, three of which were confirmed by RNA bisulfite sequencing. Using patient cells lacking the NSun2 protein, we further show that loss of cytosine-5 methylation in vault RNAs causes aberrant processing into Argonaute-associated small RNA fragments that can function as microRNAs. Thus, impaired processing of vault ncRNA may contribute to the etiology of NSun2-deficiency human disorders.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Brzezicha B., Schmidt M., Makalowska I., Jarmolowski A., Pienkowska J., Szweykowska-Kulinska Z. Identification of human tRNA:m5C methyltransferase catalysing intron-dependent m5C formation in the first position of the anticodon of the pre-tRNA Leu (CAA) Nucleic Acids Res. 2006;34:6034–6043. - PMC - PubMed
-
- Burgess D.L., Gefrides L.A., Foreman P.J., Noebels J.L. A cluster of three novel Ca2+ channel gamma subunit genes on chromosome 19q13.4: evolution and expression profile of the gamma subunit gene family. Genomics. 2001;71:339–350. - PubMed
Supplemental References
-
- Shi R., Chiang V.L. Facile means for quantifying microRNA expression by real-time PCR. Biotechniques. 2005;39:519–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
