

Learning to learn - intrinsic plasticity as a metaplasticity mechanism for memory formation
- PMID: 23871744
- PMCID: PMC3855019
- DOI: 10.1016/j.nlm.2013.07.008
Learning to learn - intrinsic plasticity as a metaplasticity mechanism for memory formation
Abstract
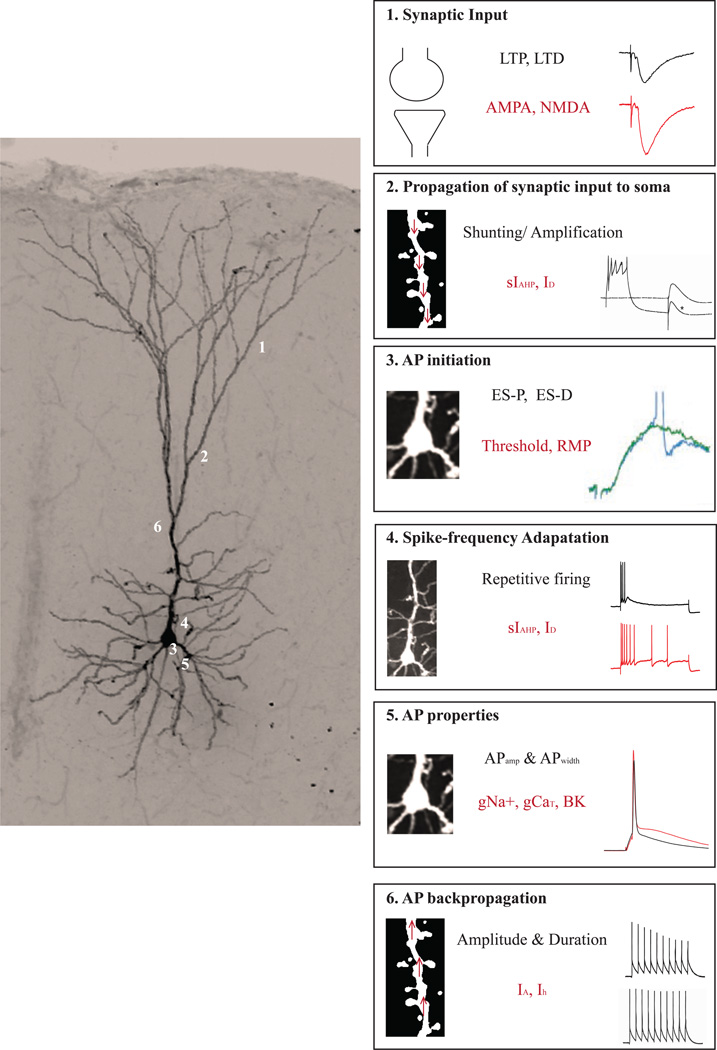
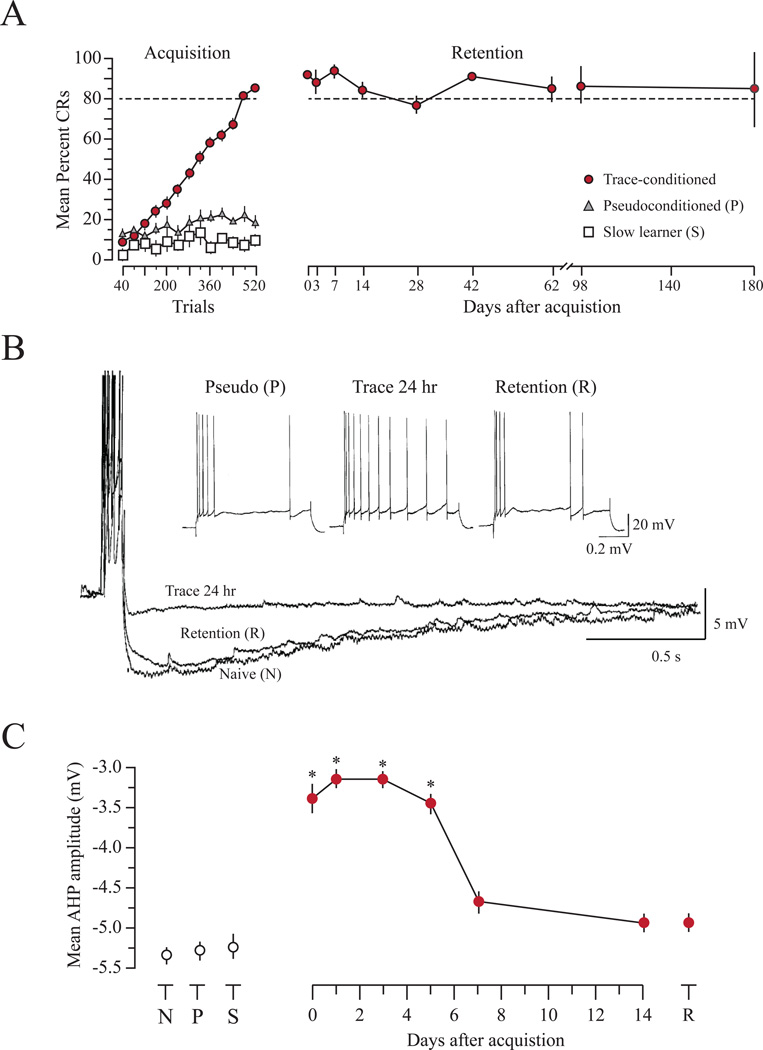
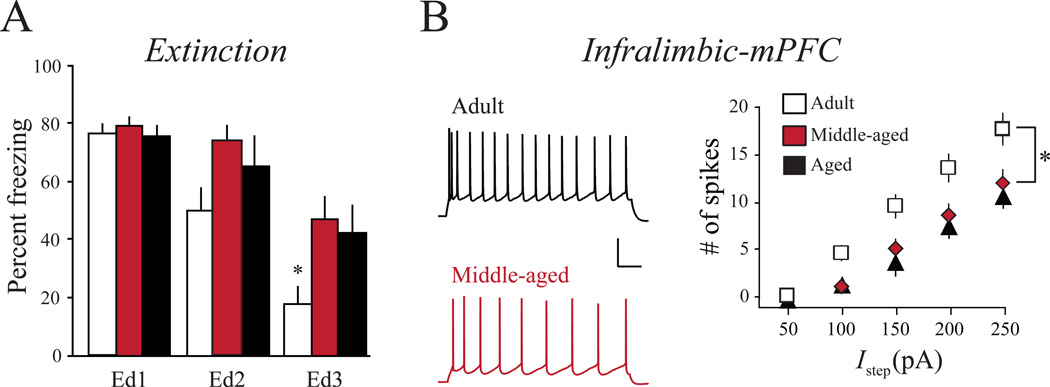
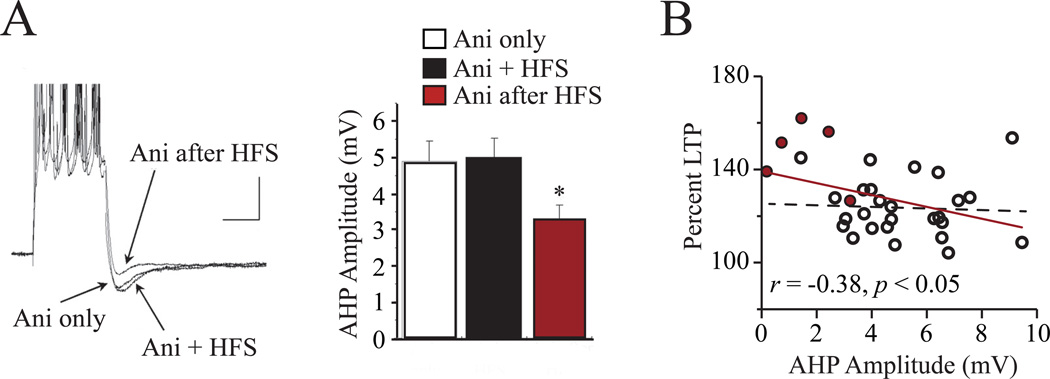
"Use it or lose it" is a popular adage often associated with use-dependent enhancement of cognitive abilities. Much research has focused on understanding exactly how the brain changes as a function of experience. Such experience-dependent plasticity involves both structural and functional alterations that contribute to adaptive behaviors, such as learning and memory, as well as maladaptive behaviors, including anxiety disorders, phobias, and posttraumatic stress disorder. With the advancing age of our population, understanding how use-dependent plasticity changes across the lifespan may also help to promote healthy brain aging. A common misconception is that such experience-dependent plasticity (e.g., associative learning) is synonymous with synaptic plasticity. Other forms of plasticity also play a critical role in shaping adaptive changes within the nervous system, including intrinsic plasticity - a change in the intrinsic excitability of a neuron. Intrinsic plasticity can result from a change in the number, distribution or activity of various ion channels located throughout the neuron. Here, we review evidence that intrinsic plasticity is an important and evolutionarily conserved neural correlate of learning. Intrinsic plasticity acts as a metaplasticity mechanism by lowering the threshold for synaptic changes. Thus, learning-related intrinsic changes can facilitate future synaptic plasticity and learning. Such intrinsic changes can impact the allocation of a memory trace within a brain structure, and when compromised, can contribute to cognitive decline during the aging process. This unique role of intrinsic excitability can provide insight into how memories are formed and, more interestingly, how neurons that participate in a memory trace are selected. Most importantly, modulation of intrinsic excitability can allow for regulation of learning ability - this can prevent or provide treatment for cognitive decline not only in patients with clinical disorders but also in the aging population.
Keywords: Aging; Intrinsic excitability; Learning; Memory; Memory allocation; Memory modulation; Metaplasticity.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Abraham WC, Bear MF. Metaplasticity: the plasticity of synaptic plasticity. Trends Neurosci. 1996;19:126–130. - PubMed
-
- Abraham WC. Metaplasticity: tuning synapses and networks for plasticity. Nat Rev Neurosci. 2008;9:387. - PubMed
-
- Alger BE, Nicoll RA. Epileptiform burst afterhyperolarization: calciumdependent potassium potential in hippocampal CA1 pyramidal cells. Science. 1980;210:1122–1124. - PubMed
-
- Alkon DL. Voltage-dependent calcium and potassium ion conductances: a contingency mechanism for an associative learning model. Science. 1979;205:810–816. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
