

A low-frequency oscillatory neural signal in humans encodes a developing decision variable
- PMID: 23872495
- PMCID: PMC3815962
- DOI: 10.1016/j.neuroimage.2013.06.085
A low-frequency oscillatory neural signal in humans encodes a developing decision variable
Abstract
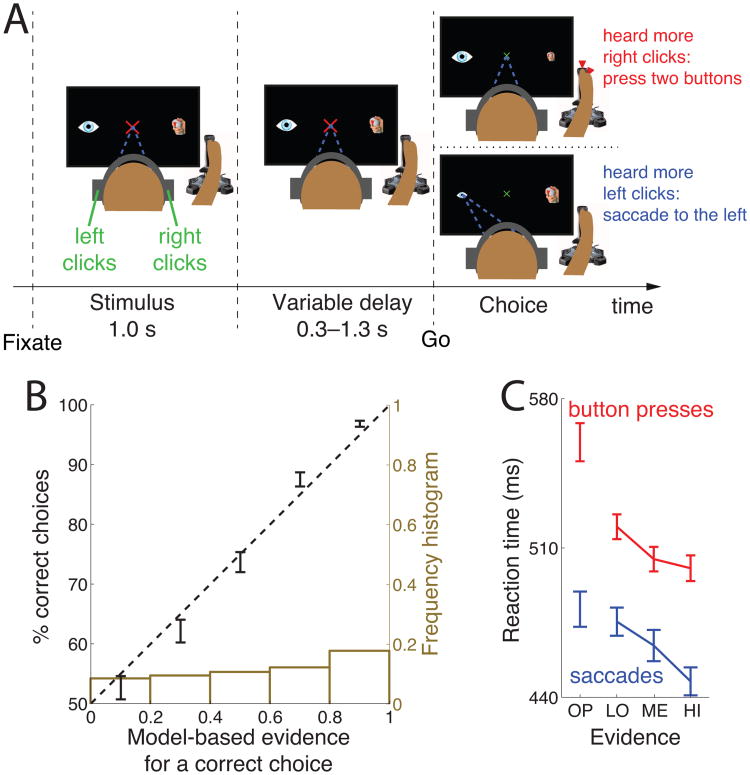
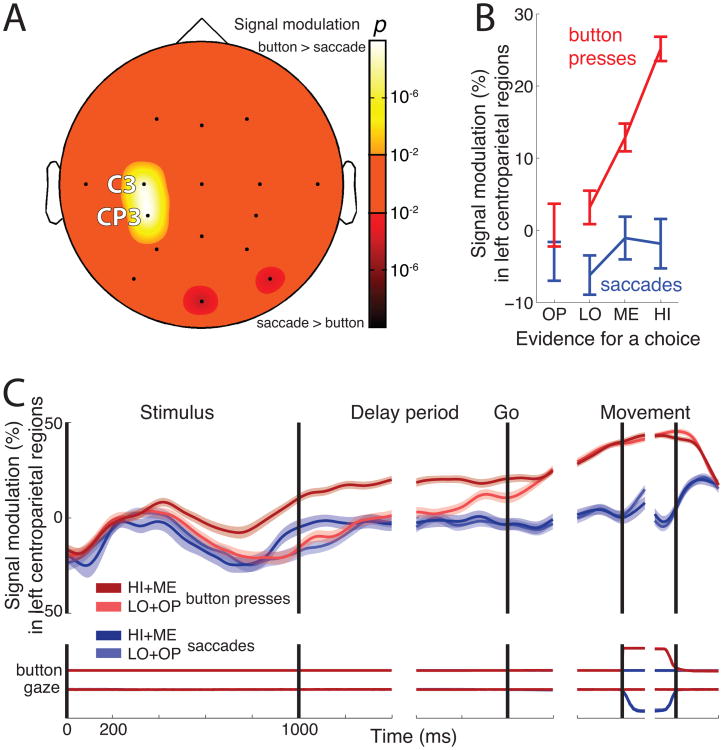
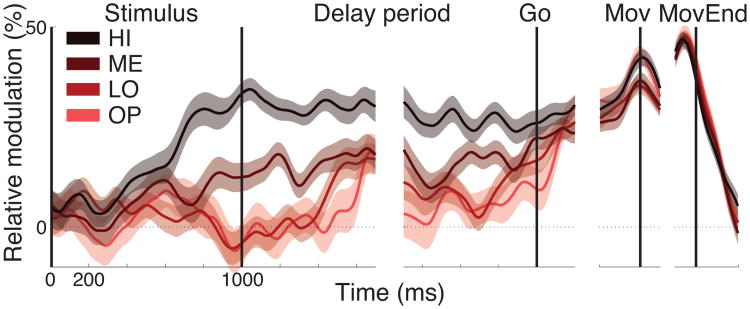
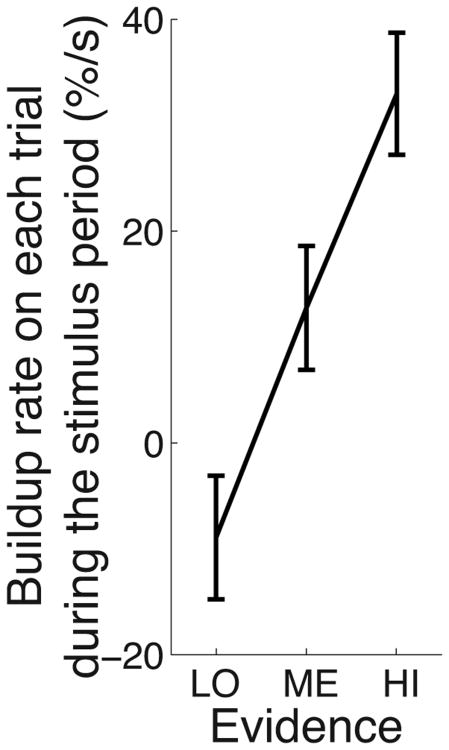
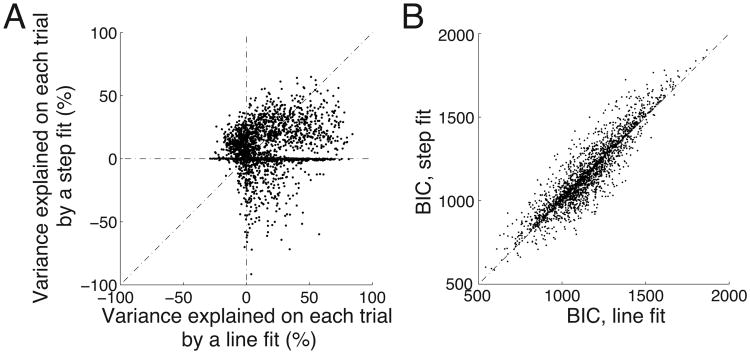
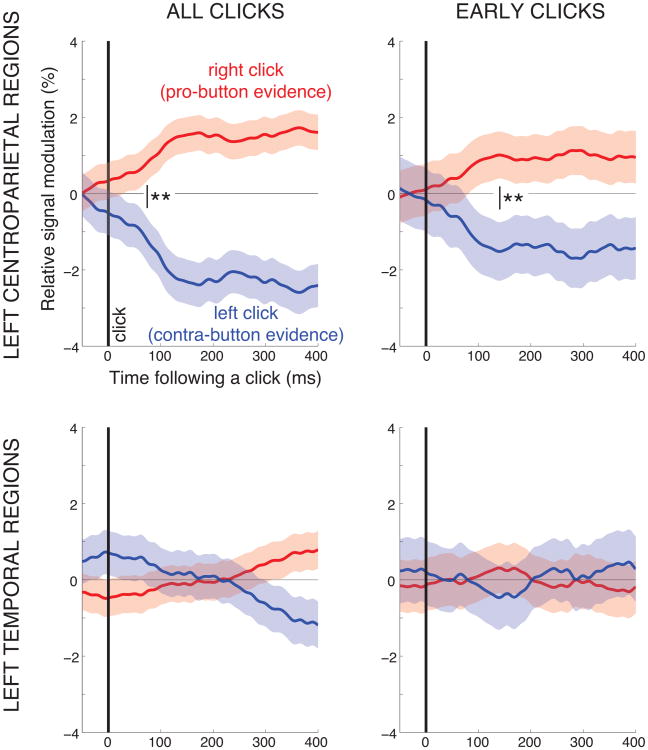
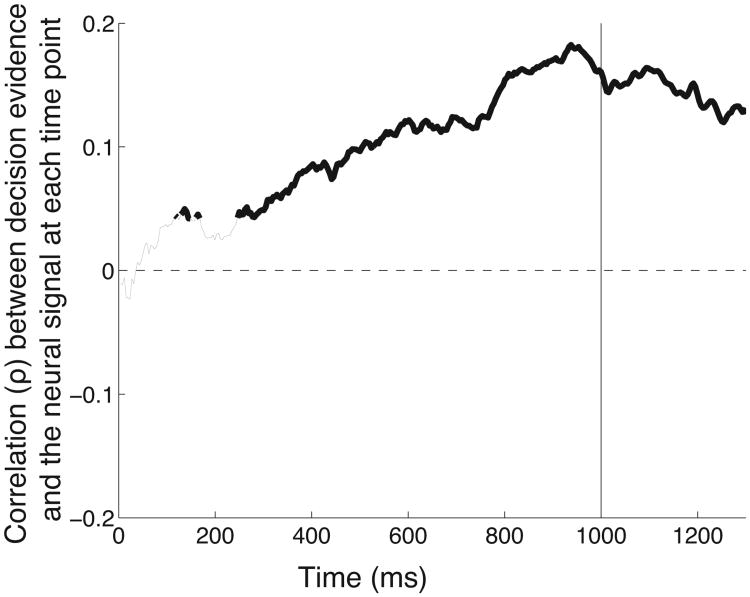
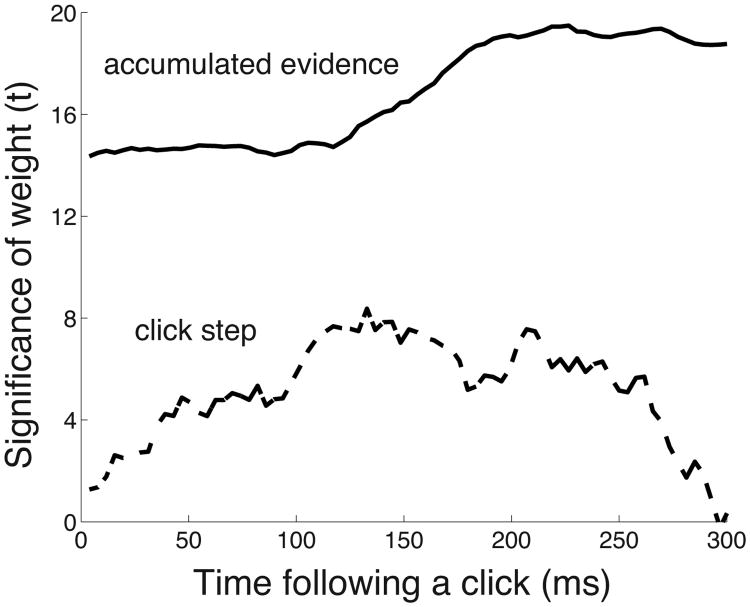
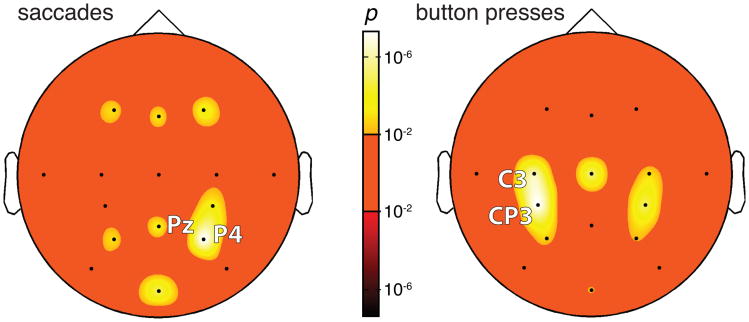
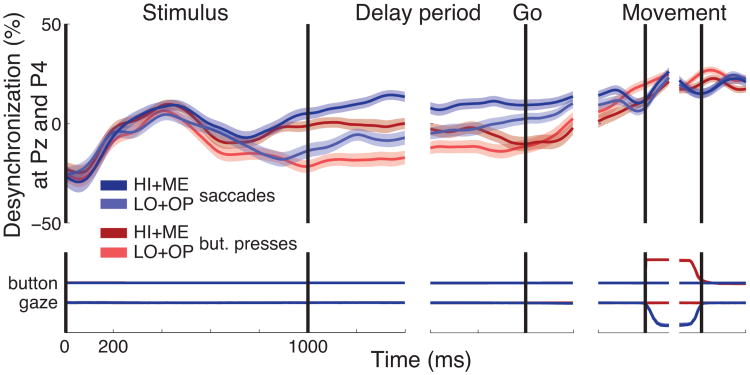
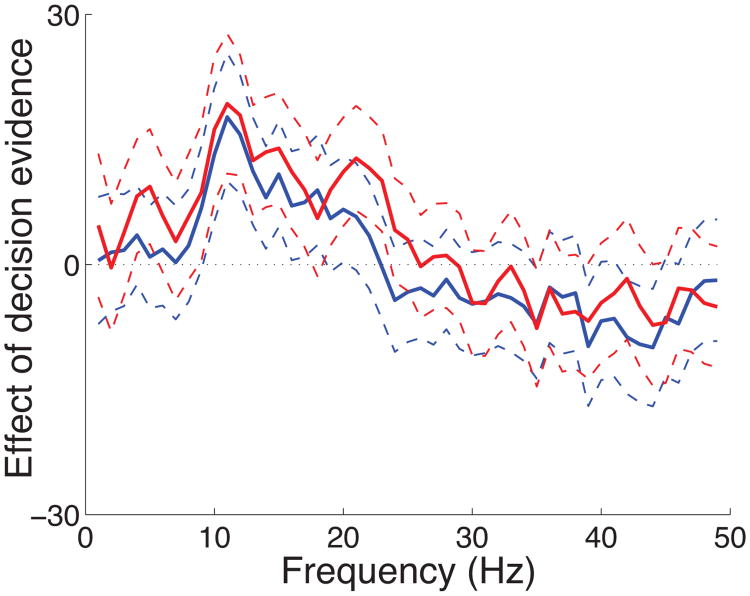
We often make decisions based on sensory evidence that is accumulated over a period of time. How the evidence for such decisions is represented in the brain and how such a neural representation is used to guide a subsequent action are questions of considerable interest to decision sciences. The neural correlates of developing perceptual decisions have been thoroughly investigated in the oculomotor system of macaques who communicated their decisions using an eye movement. It has been found that the evidence informing a decision to make an eye movement is in part accumulated within the same oculomotor circuits that signal the upcoming eye movement. Recent evidence suggests that the somatomotor system may exhibit an analogous property for choices made using a hand movement. To investigate this possibility, we engaged humans in a decision task in which they integrated discrete quanta of sensory information over a period of time and signaled their decision using a hand movement or an eye movement. The discrete form of the sensory evidence allowed us to infer the decision variable on which subjects base their decision on each trial and to assess the neural processes related to each quantum of the incoming decision evidence. We found that a low-frequency electrophysiological signal recorded over centroparietal regions strongly encodes the decision variable inferred in this task, and that it does so specifically for hand movement choices. The signal ramps up with a rate that is proportional to the decision variable, remains graded by the decision variable throughout the delay period, reaches a common peak shortly before a hand movement, and falls off shortly after the hand movement. Furthermore, the signal encodes the polarity of each evidence quantum, with a short latency, and retains the response level over time. Thus, this neural signal shows properties of evidence accumulation. These findings suggest that the decision-related effects observed in the oculomotor system of the monkey during eye movement choices may share the same basic properties with the decision-related effects in the somatomotor system of humans during hand movement choices.
© 2013 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Eye movements as a readout of sensorimotor decision processes.J Neurophysiol. 2020 Apr 1;123(4):1439-1447. doi: 10.1152/jn.00622.2019. Epub 2020 Mar 11. J Neurophysiol. 2020. PMID: 32159423 Free PMC article.
-
Correlates of perceptual learning in an oculomotor decision variable.J Neurosci. 2009 Feb 18;29(7):2136-50. doi: 10.1523/JNEUROSCI.3962-08.2009. J Neurosci. 2009. PMID: 19228966 Free PMC article.
-
Decoding go/no-go decisions from eye movements.J Vis. 2019 Feb 1;19(2):5. doi: 10.1167/19.2.5. J Vis. 2019. PMID: 30735563
-
Bridging Neural and Computational Viewpoints on Perceptual Decision-Making.Trends Neurosci. 2018 Nov;41(11):838-852. doi: 10.1016/j.tins.2018.06.005. Epub 2018 Jul 12. Trends Neurosci. 2018. PMID: 30007746 Free PMC article. Review.
-
Neural correlates of decision processes: neural and mental chronometry.Curr Opin Neurobiol. 2003 Apr;13(2):182-6. doi: 10.1016/s0959-4388(03)00039-4. Curr Opin Neurobiol. 2003. PMID: 12744971 Review.
Cited by
-
Within-subject reaction time variability: Role of cortical networks and underlying neurophysiological mechanisms.Neuroimage. 2021 Aug 15;237:118127. doi: 10.1016/j.neuroimage.2021.118127. Epub 2021 May 4. Neuroimage. 2021. PMID: 33957232 Free PMC article.
-
Beta oscillations reflect supramodal information during perceptual judgment.Proc Natl Acad Sci U S A. 2017 Dec 26;114(52):13810-13815. doi: 10.1073/pnas.1714633115. Epub 2017 Dec 11. Proc Natl Acad Sci U S A. 2017. PMID: 29229820 Free PMC article.
-
Sensorimotor faculties bias perceptual decision-making.bioRxiv [Preprint]. 2024 May 16:2024.05.14.594024. doi: 10.1101/2024.05.14.594024. bioRxiv. 2024. PMID: 38798638 Free PMC article. Preprint.
-
State-dependent architecture of thalamic reticular subnetworks.Cell. 2014 Aug 14;158(4):808-821. doi: 10.1016/j.cell.2014.06.025. Cell. 2014. PMID: 25126786 Free PMC article.
-
Graded decisions in the human brain.Nat Commun. 2024 May 21;15(1):4308. doi: 10.1038/s41467-024-48342-w. Nat Commun. 2024. PMID: 38773117 Free PMC article.
References
-
- Anderson ML. Embodied cognition: A field guide. Artificial Intelligence. 2003;149:91–130.
-
- Brooks R. Intelligence without representation. Artificial Intelligence. 1991;47:139–159.
-
- Brunton BW, Botvinick MM, Brody CD. Rats and humans can optimally accumulate evidence for decision-making. Science. 2013;340:95–98. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
