

Involvement of amygdalar protein kinase A, but not calcium/calmodulin-dependent protein kinase II, in the reconsolidation of cocaine-related contextual memories in rats
- PMID: 23873418
- PMCID: PMC3852194
- DOI: 10.1007/s00213-013-3203-9
Involvement of amygdalar protein kinase A, but not calcium/calmodulin-dependent protein kinase II, in the reconsolidation of cocaine-related contextual memories in rats
Abstract
Rationale: Contextual control over drug relapse depends on the successful reconsolidation and retention of context-response-cocaine associations in long-term memory stores. The basolateral amygdala (BLA) plays a critical role in cocaine memory reconsolidation and subsequent drug context-induced cocaine-seeking behavior; however, less is known about the cellular mechanisms of this phenomenon.
Objectives: The present study evaluated the hypothesis that protein kinase A (PKA) and calcium/calmodulin-dependent protein kinase II (CaMKII) activation in the BLA is necessary for the reconsolidation of context-response-cocaine memories that promote subsequent drug context-induced cocaine-seeking behavior.
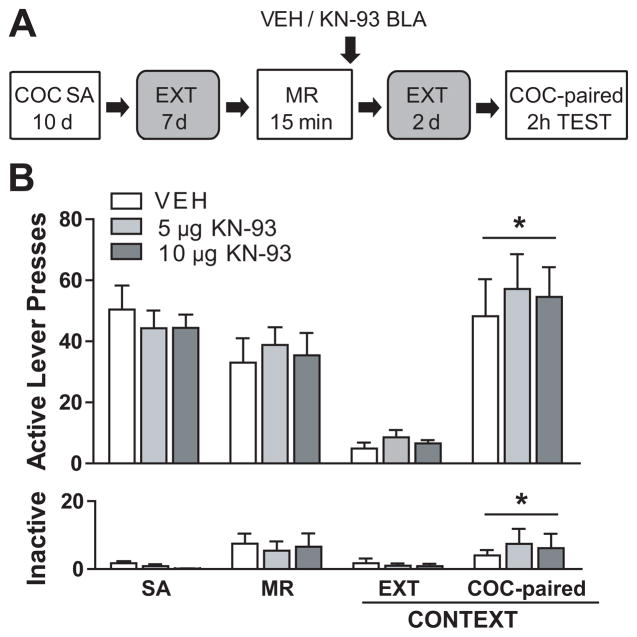
Methods: Rats were trained to lever-press for cocaine infusions in a distinct context, followed by extinction training in a different context. Rats were then briefly re-exposed to the previously cocaine-paired context or an unpaired context in order to reactivate cocaine-related contextual memories and initiate their reconsolidation or to provide a similar behavioral experience without explicit cocaine-related memory reactivation, respectively. Immediately after this session, rats received bilateral microinfusions of vehicle, the PKA inhibitor, Rp-adenosine 3',5'-cyclic monophosphorothioate triethylammonium salt (Rp-cAMPS), or the CaMKII inhibitor, KN-93, into the BLA or the posterior caudate putamen (anatomical control region). Rats were then tested for cocaine-seeking behavior (responses on the previously cocaine-paired lever) in the cocaine-paired context and the extinction context.
Results: Intra-BLA infusion of Rp-cAMPS, but not KN-93, following cocaine memory reconsolidation impaired subsequent cocaine-seeking behavior in a dose-dependent, site-specific, and memory reactivation-dependent fashion.
Conclusions: PKA, but not CaMKII, activation in the BLA is critical for cocaine memory re-stabilization processes that facilitate subsequent drug context-induced instrumental cocaine-seeking behavior.
Conflict of interest statement
The authors have no conflicts of interest to disclose.
Figures




References
-
- Arnsten ART, Ramos BP, Birnbaum SG, Taylor JR. Protein kinase A as a therapeutic target for memory disorders: rationale and challenges. Trends Mol Med. 2005;11:121–8. - PubMed
-
- Ben Mamou C, Gamache K, Nader K. NMDA receptors are critical for unleashing consolidated auditory fear memories. Nat Neurosci. 2006;9:1237–1239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
