

Inferring nonlinear neuronal computation based on physiologically plausible inputs
- PMID: 23874185
- PMCID: PMC3715434
- DOI: 10.1371/journal.pcbi.1003143
Inferring nonlinear neuronal computation based on physiologically plausible inputs
Abstract
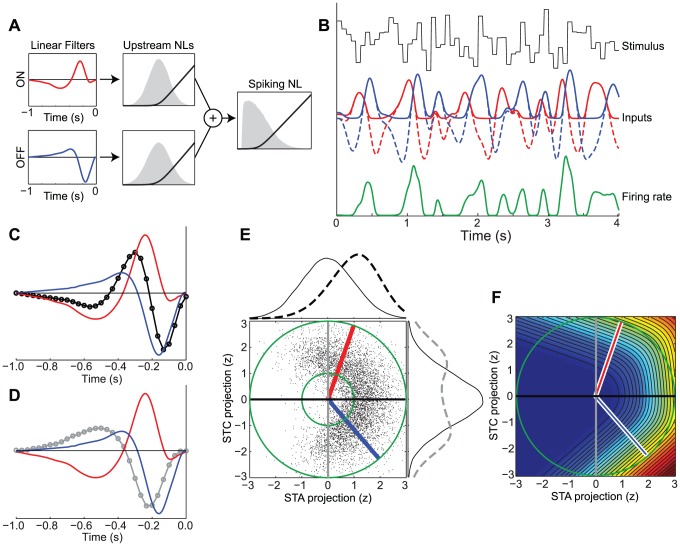
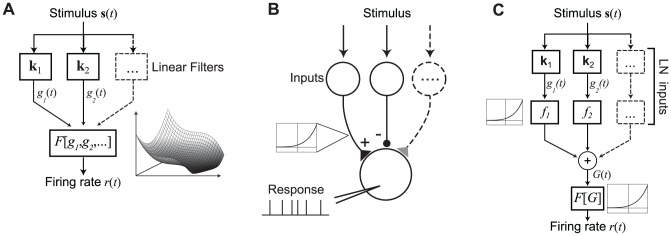
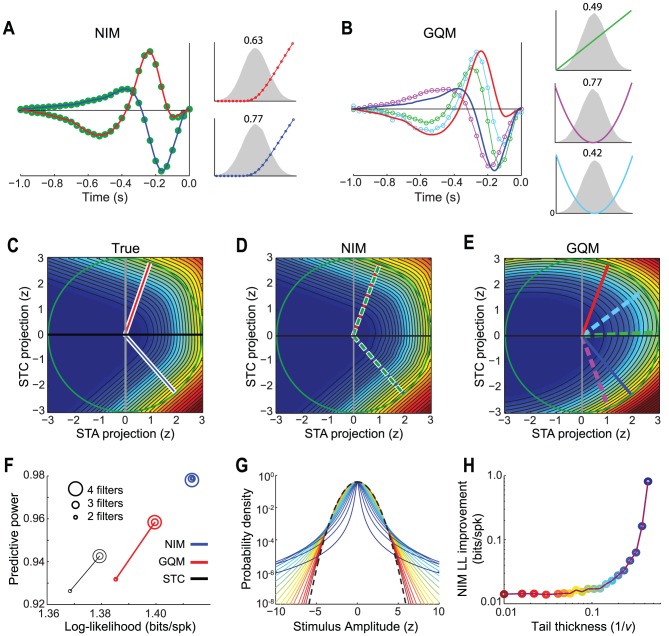
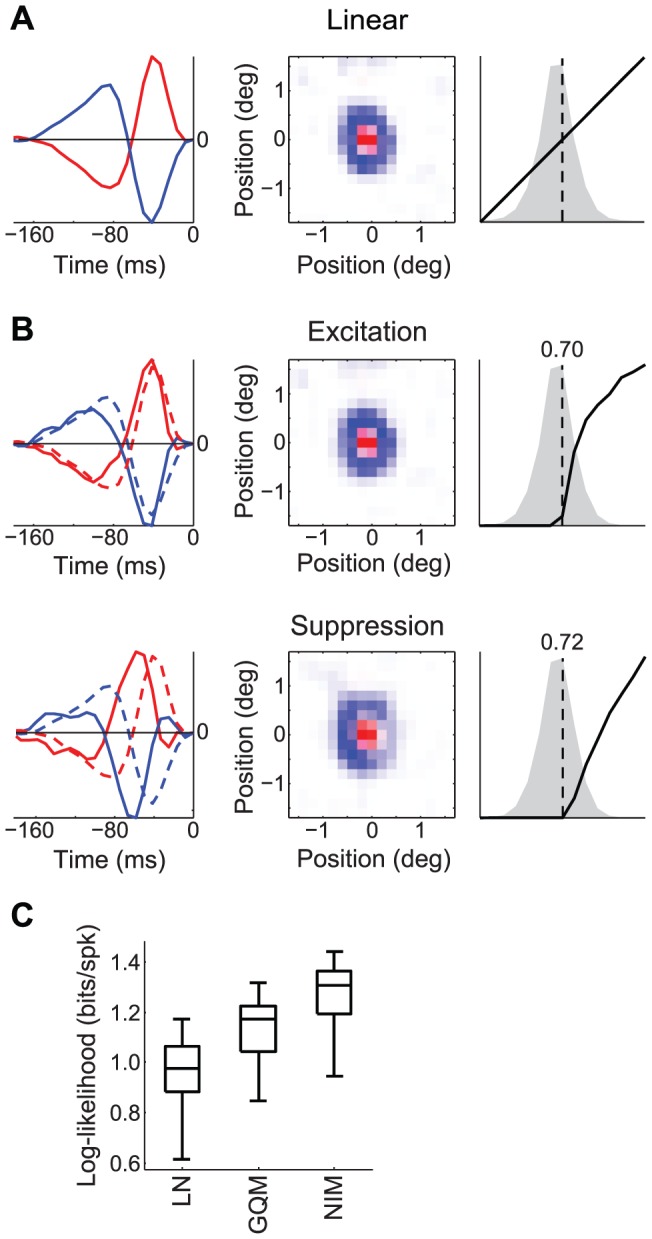
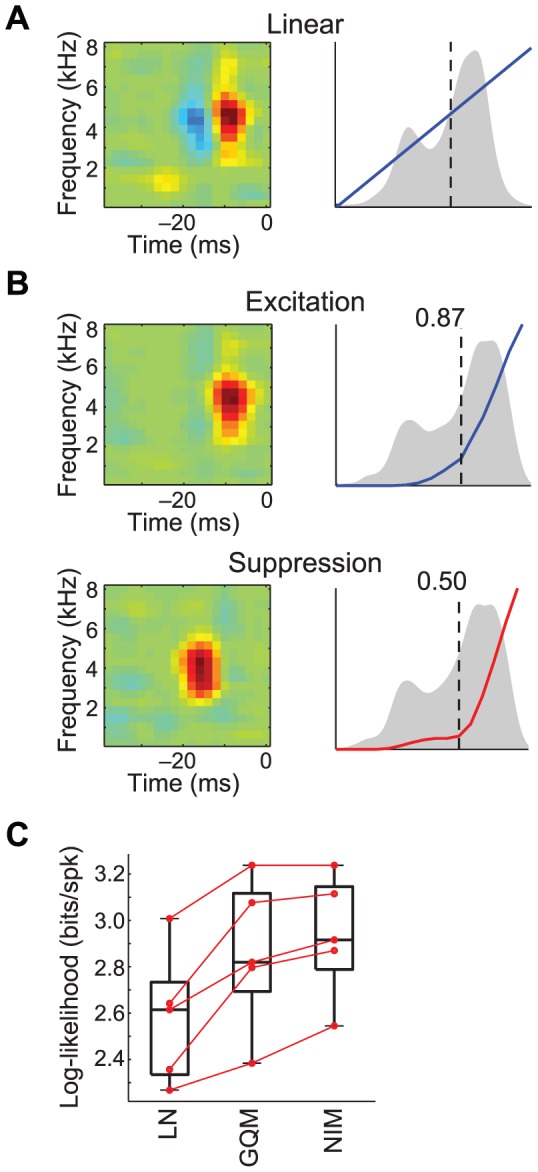
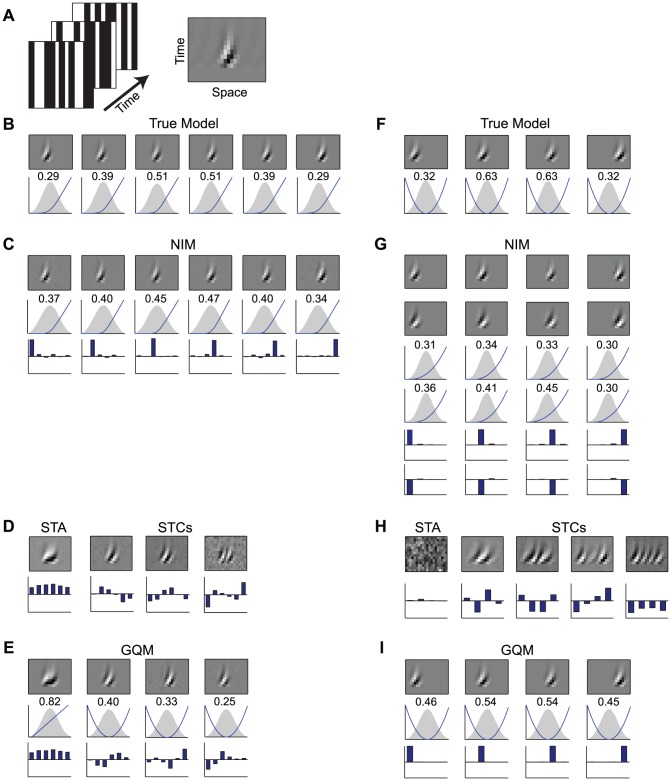
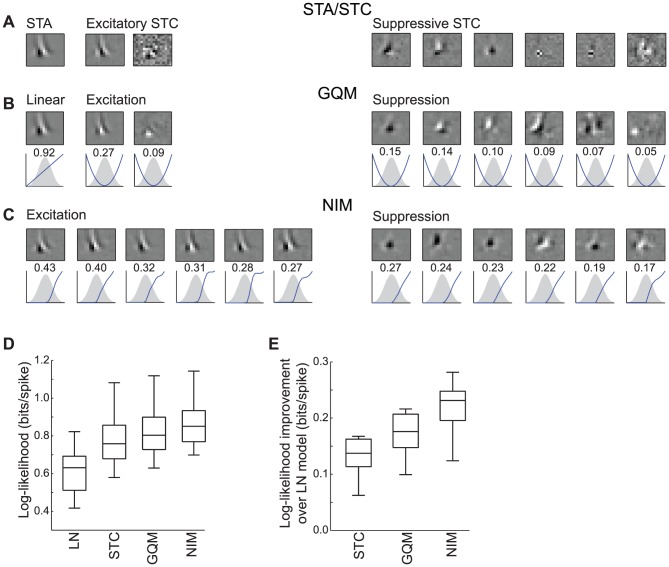
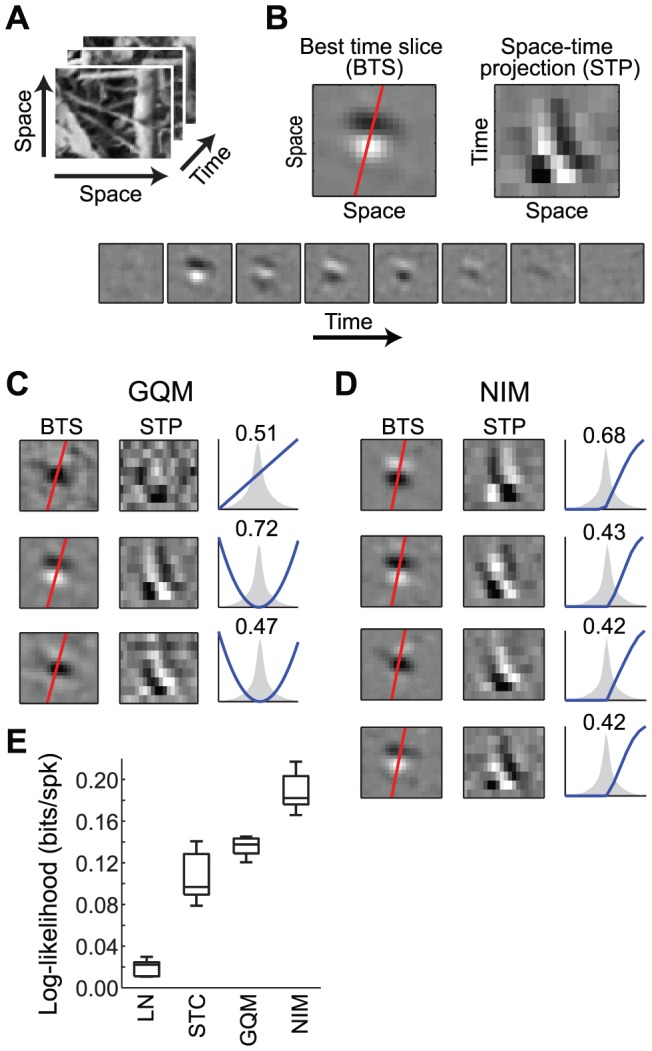
The computation represented by a sensory neuron's response to stimuli is constructed from an array of physiological processes both belonging to that neuron and inherited from its inputs. Although many of these physiological processes are known to be nonlinear, linear approximations are commonly used to describe the stimulus selectivity of sensory neurons (i.e., linear receptive fields). Here we present an approach for modeling sensory processing, termed the Nonlinear Input Model (NIM), which is based on the hypothesis that the dominant nonlinearities imposed by physiological mechanisms arise from rectification of a neuron's inputs. Incorporating such 'upstream nonlinearities' within the standard linear-nonlinear (LN) cascade modeling structure implicitly allows for the identification of multiple stimulus features driving a neuron's response, which become directly interpretable as either excitatory or inhibitory. Because its form is analogous to an integrate-and-fire neuron receiving excitatory and inhibitory inputs, model fitting can be guided by prior knowledge about the inputs to a given neuron, and elements of the resulting model can often result in specific physiological predictions. Furthermore, by providing an explicit probabilistic model with a relatively simple nonlinear structure, its parameters can be efficiently optimized and appropriately regularized. Parameter estimation is robust and efficient even with large numbers of model components and in the context of high-dimensional stimuli with complex statistical structure (e.g. natural stimuli). We describe detailed methods for estimating the model parameters, and illustrate the advantages of the NIM using a range of example sensory neurons in the visual and auditory systems. We thus present a modeling framework that can capture a broad range of nonlinear response functions while providing physiologically interpretable descriptions of neural computation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Riesenhuber M, Poggio T (1999) Hierarchical models of object recognition in cortex. Nat Neurosci 2: 1019–1025. - PubMed
-
- Brincat SL, Connor CE (2004) Underlying principles of visual shape selectivity in posterior inferotemporal cortex. Nat Neurosci 7: 880–886. - PubMed
-
- DiCarlo JJ, Cox DD (2007) Untangling invariant object recognition. Trends Cogn Sci 11: 333–341. - PubMed
-
- Rust NC, Stocker AA (2010) Ambiguity and invariance: two fundamental challenges for visual processing. Curr Opin Neurobiol 20: 382–388. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
