

Gata4 is required for formation of the genital ridge in mice
- PMID: 23874227
- PMCID: PMC3708810
- DOI: 10.1371/journal.pgen.1003629
Gata4 is required for formation of the genital ridge in mice
Abstract
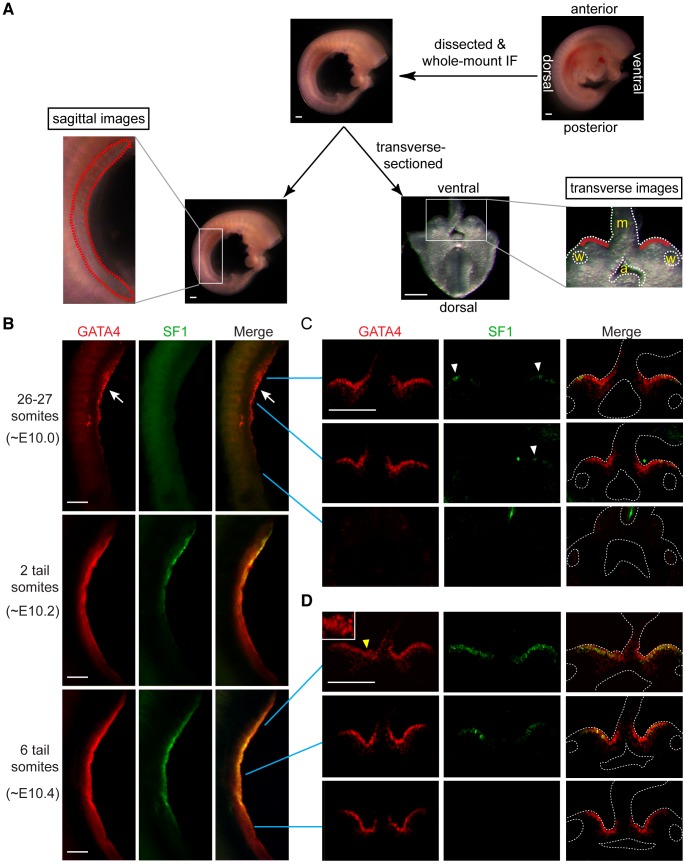
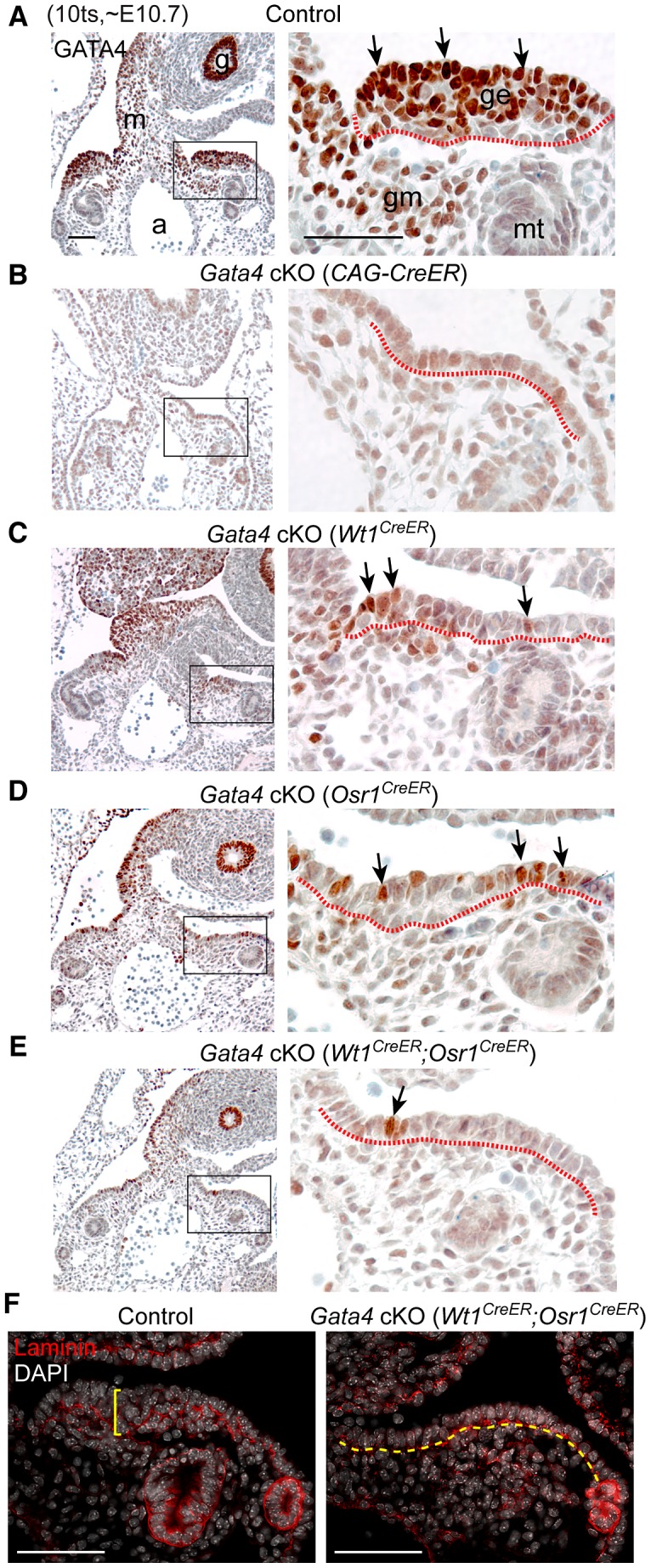
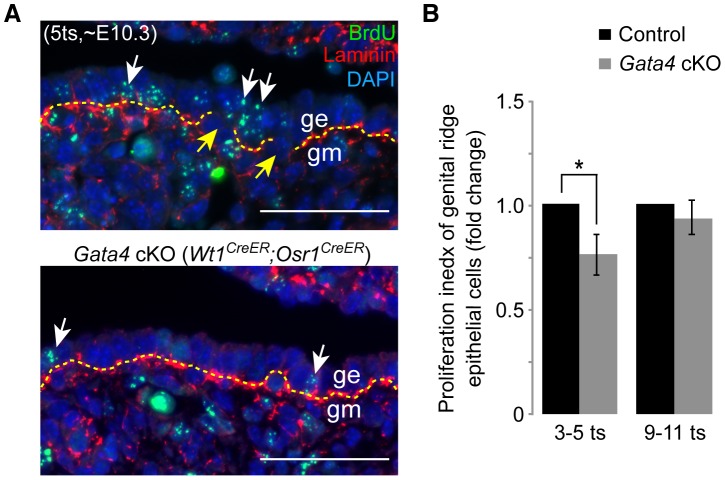
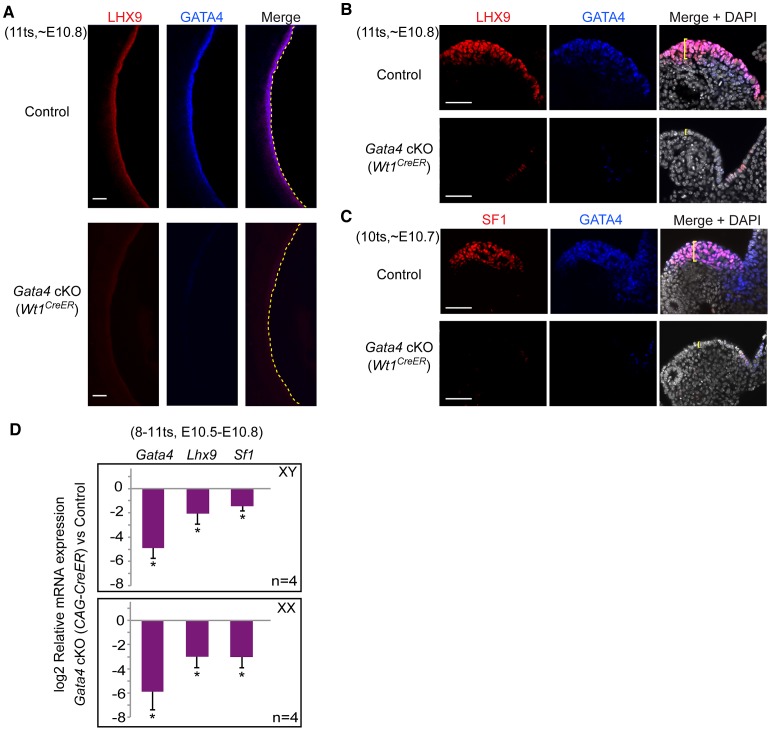
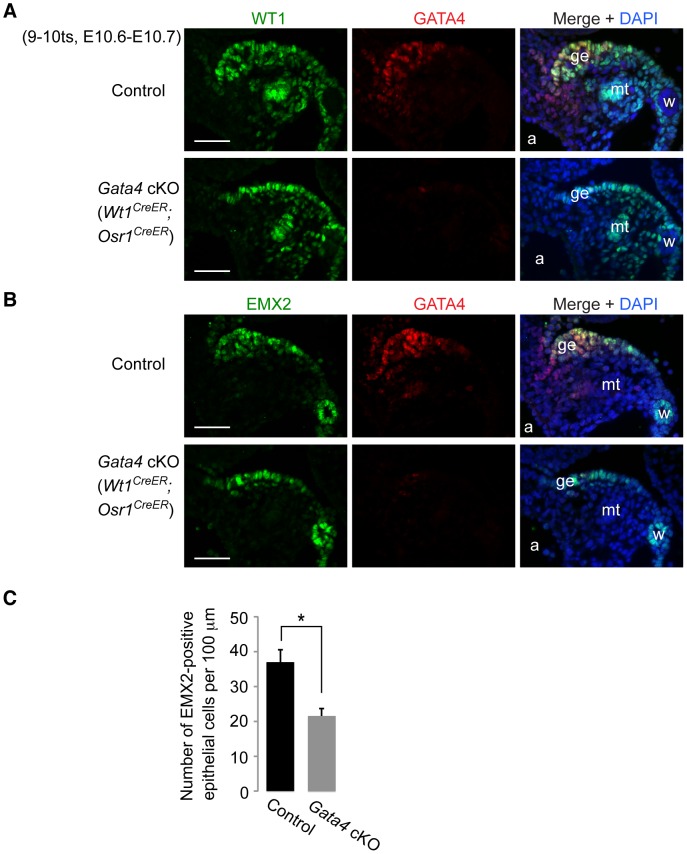
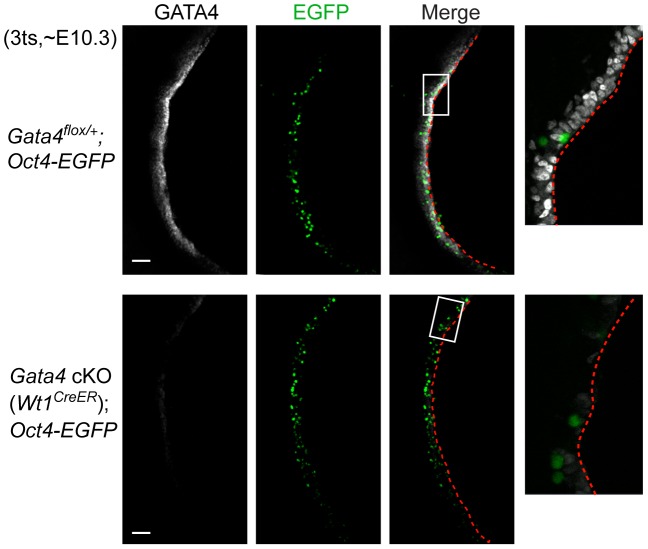
In mammals, both testis and ovary arise from a sexually undifferentiated precursor, the genital ridge, which first appears during mid-gestation as a thickening of the coelomic epithelium on the ventromedial surface of the mesonephros. At least four genes (Lhx9, Sf1, Wt1, and Emx2) have been demonstrated to be required for subsequent growth and maintenance of the genital ridge. However, no gene has been shown to be required for the initial thickening of the coelomic epithelium during genital ridge formation. We report that the transcription factor GATA4 is expressed in the coelomic epithelium of the genital ridge, progressing in an anterior-to-posterior (A-P) direction, immediately preceding an A-P wave of epithelial thickening. Mouse embryos conditionally deficient in Gata4 show no signs of gonadal initiation, as their coelomic epithelium remains a morphologically undifferentiated monolayer. The failure of genital ridge formation in Gata4-deficient embryos is corroborated by the absence of the early gonadal markers LHX9 and SF1. Our data indicate that GATA4 is required to initiate formation of the genital ridge in both XX and XY fetuses, prior to its previously reported role in testicular differentiation of the XY gonad.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
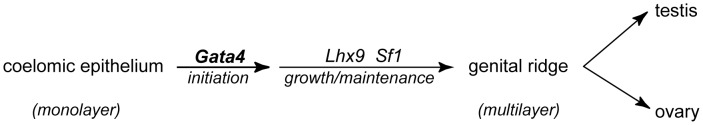
References
-
- Brambell FWR (1927) The development and morphology of the gonads of the mouse - Part I The morphogenesis of the indifferent gonad and of the ovary. Proc R Soc Lond B Biol Sci 101: 391–409.
-
- Gropp A, Ohno S (1966) The presence of a common embryonic blastema for ovarian and testicular parenchymal (follicular, interstitial and tubular) cells in cattle Bos taurus . Z Zellforsch Mikrosk Anat 74: 505–528. - PubMed
-
- Pelliniemi LJ (1975) Ultrastructure of gonadal ridge in male and female pig embryos. Anat Embryol (Berl) 147: 20–34. - PubMed
-
- Wartenberg H, Kinsky I, Viebahn C, Schmolke C (1991) Fine structural characteristics of testicular cord formation in the developing rabbit gonad. J Electron Microsc Tech 19: 133–157. - PubMed
-
- Karl J, Capel B (1998) Sertoli cells of the mouse testis originate from the coelomic epithelium. Dev Biol 203: 323–333. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
