

Morphological and molecular characterization of dietary-induced pseudo-albinism during post-embryonic development of Solea senegalensis (Kaup, 1858)
- PMID: 23874785
- PMCID: PMC3712922
- DOI: 10.1371/journal.pone.0068844
Morphological and molecular characterization of dietary-induced pseudo-albinism during post-embryonic development of Solea senegalensis (Kaup, 1858)
Abstract
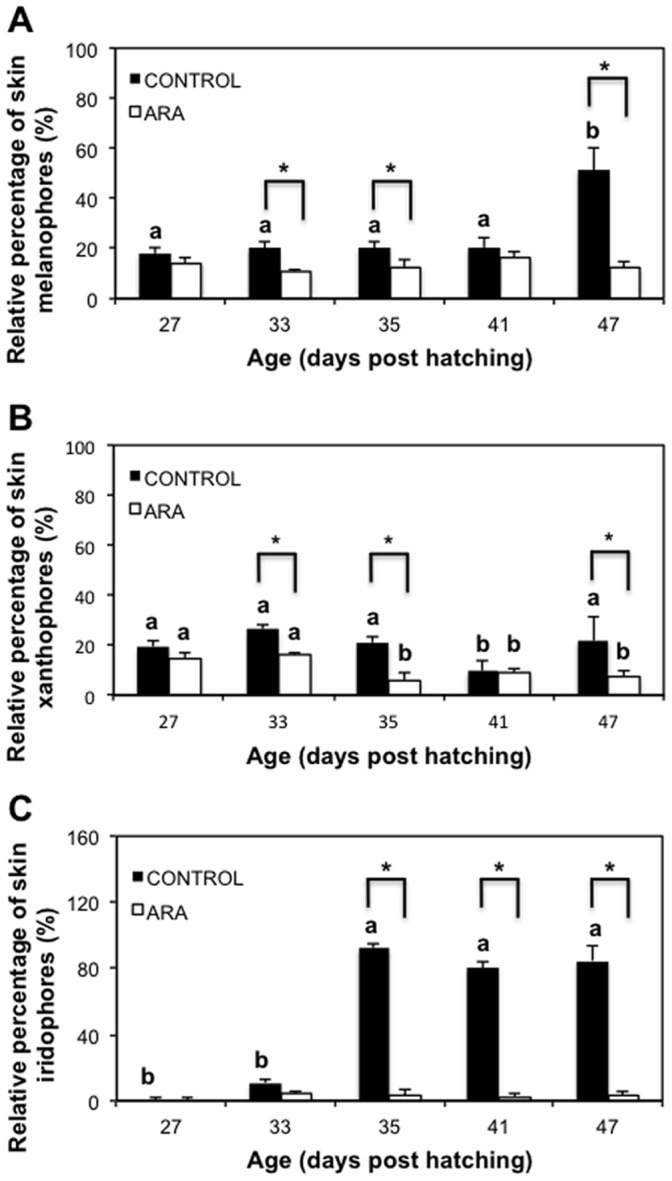
The appearance of the pseudo-albino phenotype was investigated in developing Senegalese sole (Solea senegalensis, Kaup 1858) larvae at morphological and molecular levels. In order to induce the development of pseudo-albinos, Senegalese sole larvae were fed Artemia enriched with high levels of arachidonic acid (ARA). The development of their skin pigmentation was compared to that of a control group fed Artemia enriched with a reference commercial product. The relative amount of skin melanophores, xanthophores and iridophores revealed that larval pigmentation developed similarly in both groups. However, results from different relative proportions, allocation patterns, shapes and sizes of skin chromatophores revealed changes in the pigmentation pattern between ARA and control groups from 33 days post hatching onwards. The new populations of chromatophores that should appear at post-metamorphosis were not formed in the ARA group. Further, spatial patterns of distribution between the already present larval xanthophores and melanophores were suggestive of short-range interaction that seemed to be implicated in the degradation of these chromatophores, leading to the appearance of the pseudo-albino phenotype. The expression profile of several key pigmentation-related genes revealed that melanophore development was promoted in pseudo-albinos without a sufficient degree of terminal differentiation, thus preventing melanogenesis. Present results suggest the potential roles of asip1 and slc24a5 genes on the down-regulation of trp1 expression, leading to defects in melanin production. Moreover, gene expression data supports the involvement of pax3, mitf and asip1 genes in the developmental disruption of the new post-metamorphic populations of melanophores, xanthophores and iridophores.
Conflict of interest statement
Figures







References
-
- Montoliu L, Oetting WS, Bennett DC (2011) Color Genes. European Society for Pigment Cell Research. Available: http://www.espcr.org/micemut Accessed 2013 Jun 10.
-
- Fujii R (2000) The Regulation of Motile Activity in Fish Chromatophores. Pigment Cell Res 13: 300–319. - PubMed
-
- Lister JA (2002) Development of pigment cells in the zebrafish embryo. Micros Res Tech 58: 435–441. - PubMed
-
- Mellgrem EM, Johnson SL (2002) The evolution of morphological complexity in zebrafish stripes. Trends Genet 18: 128–134. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
