

Low-molecular-weight DNA replication intermediates in Escherichia coli: mechanism of formation and strand specificity
- PMID: 23876705
- PMCID: PMC3812275
- DOI: 10.1016/j.jmb.2013.07.021
Low-molecular-weight DNA replication intermediates in Escherichia coli: mechanism of formation and strand specificity
Abstract
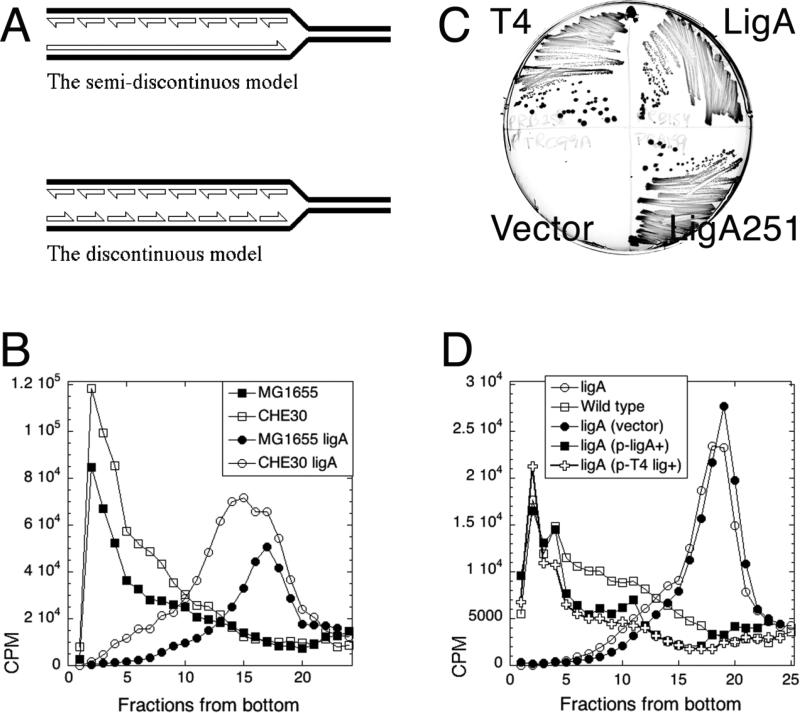
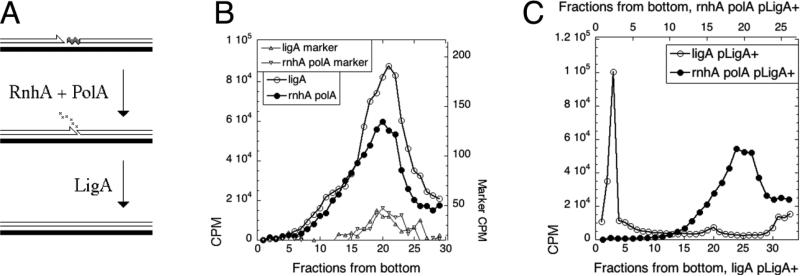
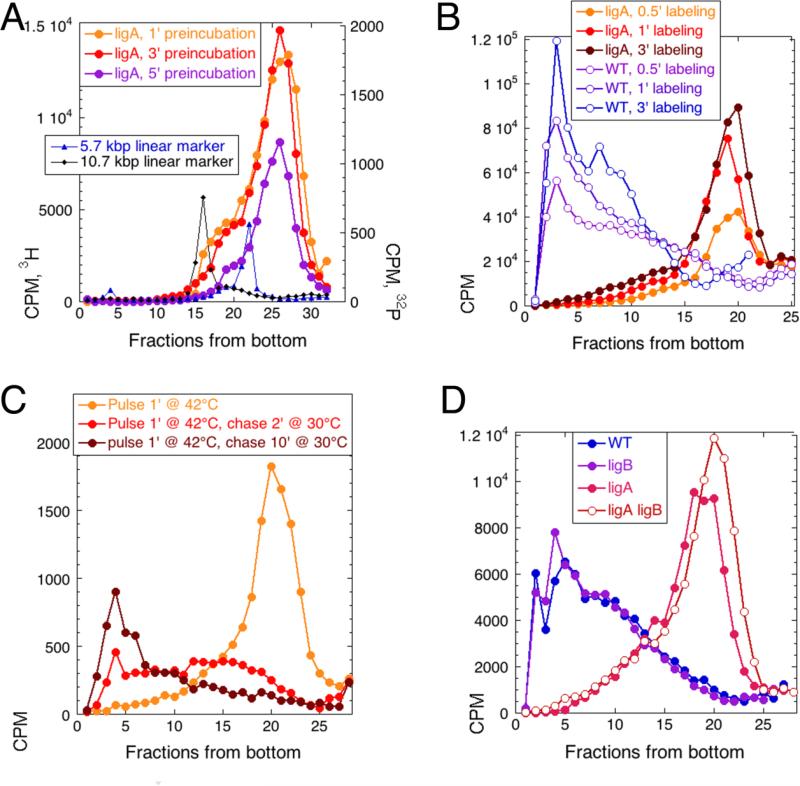
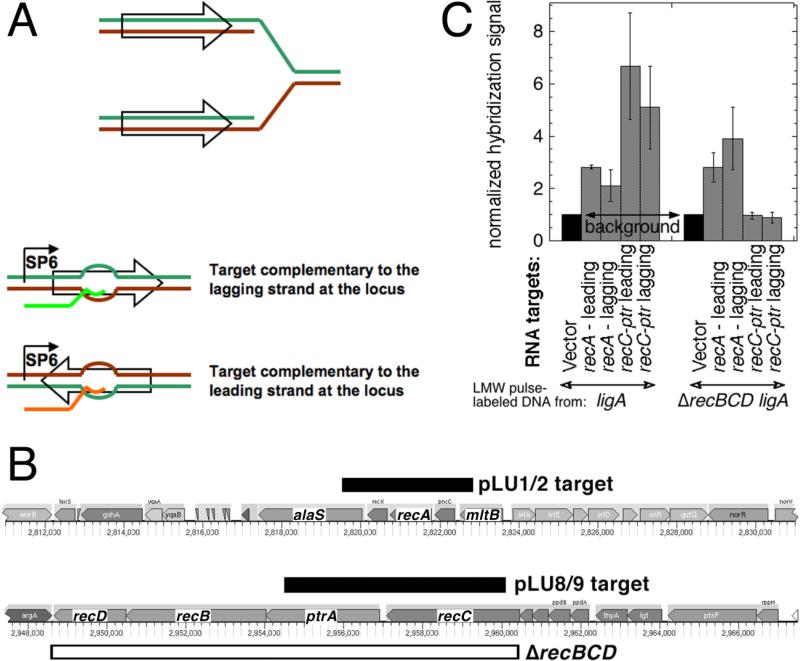
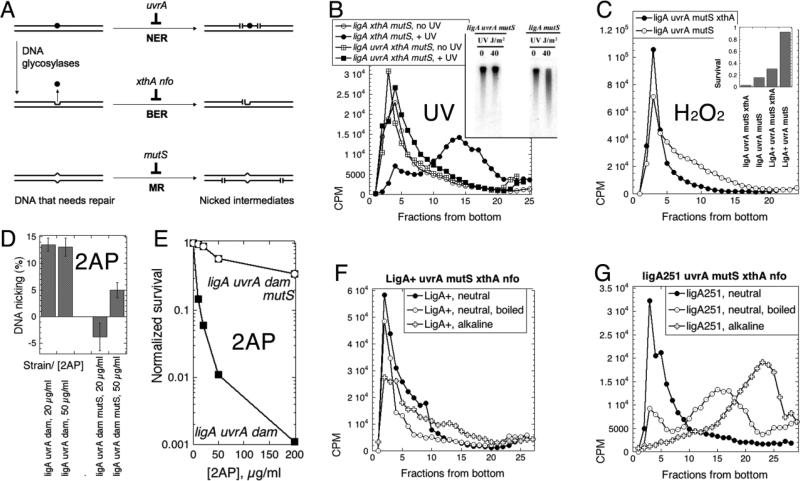
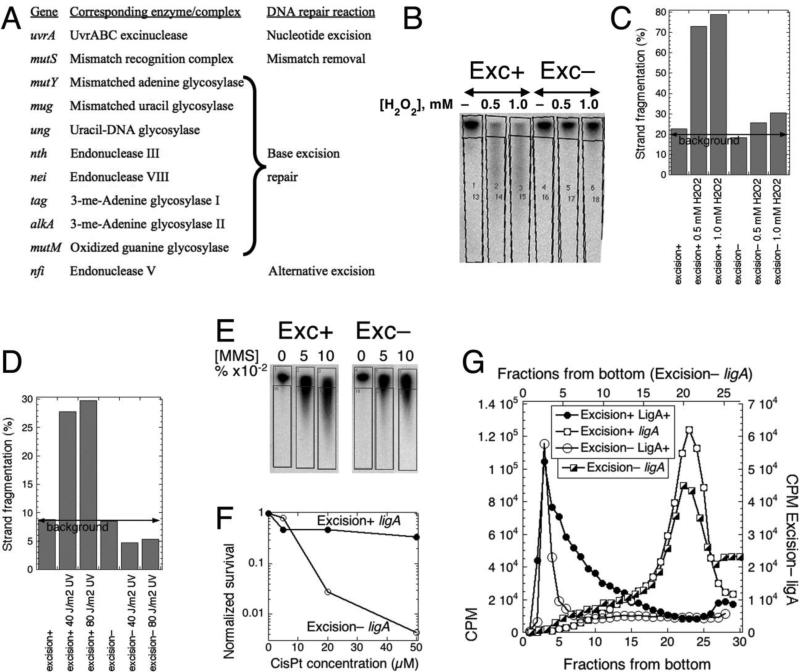
Chromosomal DNA replication intermediates, revealed in ligase-deficient conditions in vivo, are of low molecular weight (LMW) independently of the organism, suggesting discontinuous replication of both the leading and the lagging DNA strands. Yet, in vitro experiments with purified enzymes replicating sigma-structured substrates show continuous synthesis of the leading DNA strand in complete absence of ligase, supporting the textbook model of semi-discontinuous DNA replication. The discrepancy between the in vivo and in vitro results is rationalized by proposing that various excision repair events nick continuously synthesized leading strands after synthesis, producing the observed LMW intermediates. Here, we show that, in an Escherichia coli ligase-deficient strain with all known excision repair pathways inactivated, new DNA is still synthesized discontinuously. Furthermore, hybridization to strand-specific targets demonstrates that the LMW replication intermediates come from both the lagging and the leading strands. These results support the model of discontinuous leading strand synthesis in E. coli.
Keywords: BER; EDTA; HMW; IMW; LMW; MR; NER; Okazaki fragments; base excision repair; ethylenediaminetetraacetic acid; excision repair; high molecular weight; intermediate molecular weight; ligA mutants; low molecular weight; mismatch repair; nucleotide excision repair; pulse labeling; strand-specific hybridization.
© 2013.
Figures
References
-
- Amado L, Kuzminov A. The replication intermediates in Escherichia coli are not the product of DNA processing or uracil excision. J. Biol. Chem. 2006;281:22635–22646. - PubMed
-
- Gottesman MM, Hicks ML, Gellert M. Genetics and function of DNA ligase in Escherichia coli. J. Mol. Biol. 1973;77:531–547. - PubMed
-
- Horiuchi T, Sato T, Nagata T. DNA degradation in an amber mutant of Escherichia coli K12 affecting DNA ligase and viability. J. Mol. Biol. 1975;95:271–287. - PubMed
-
- Konrad EB, Modrich P, Lehman IR. Genetic and enzymatic characatrization of a conditional lethal mutant of Escherichia coli K12 with a temperature-sensitive DNA ligase. J. Mol. Biol. 1973;77:519–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
