

Disruption of TAB1/p38α interaction using a cell-permeable peptide limits myocardial ischemia/reperfusion injury
- PMID: 23877036
- PMCID: PMC3776642
- DOI: 10.1038/mt.2013.90
Disruption of TAB1/p38α interaction using a cell-permeable peptide limits myocardial ischemia/reperfusion injury
Abstract
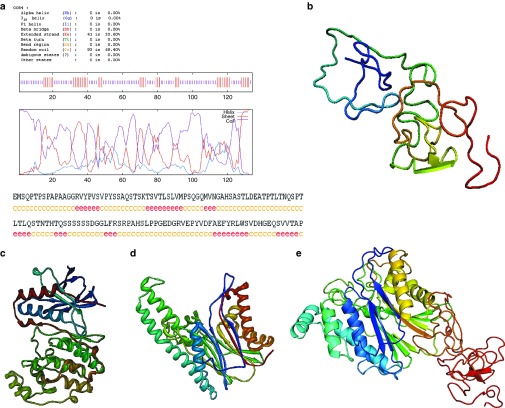
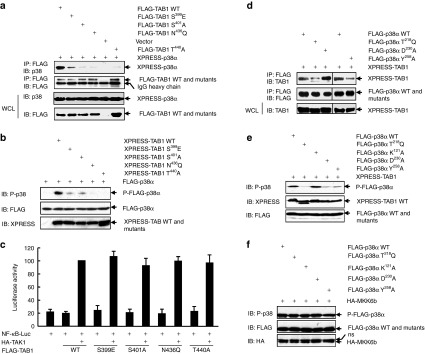
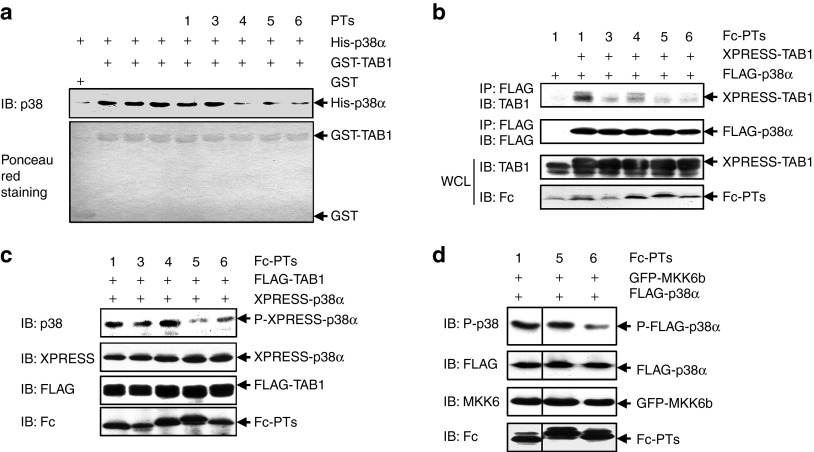
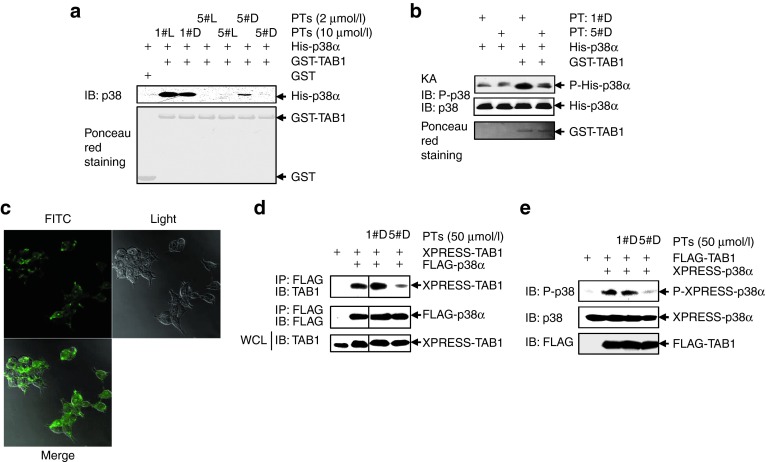
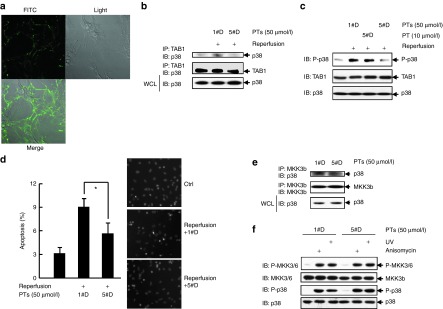
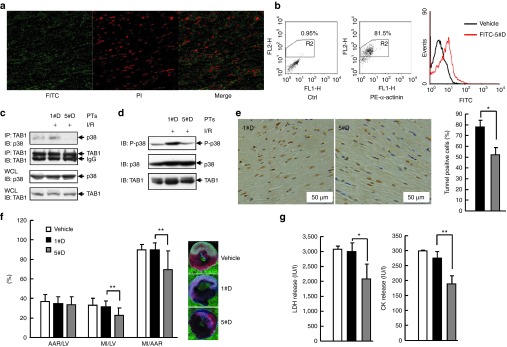
Targeting the adaptor protein (transforming growth factor-β (TGF-β)-activated protein kinase 1 (TAK1)-binding protein 1) (TAB1)-mediated non-canonical activation of p38α to limit ischemia/reperfusion (I/R) injury after an acute myocardial infarction seems to be attractive since TAB1/p38α interaction occurs specifically in very limited circumstances and possesses unique structural basis. However, so far no TAB1/p38α interaction inhibitor has been reported due to the limited knowledge about the interfaces. In this study, we sought to identify key amino acids essential for the unique mode of interaction with computer-guided molecular simulations and molecular docking. After validation of the predicted three-dimensional (3-D) structure of TAB1/p38α complex, we designed several peptides and evaluated whether they could block TAB1/p38α interaction with selectivity. We found that a cell-permeable peptide worked as a selective TAB1/p38α interaction inhibitor and decreased myocardial I/R injury. To our knowledge, this is the first TAB1/p38α interaction inhibitor.
Figures

References
-
- Denise Martin E, De Nicola GF, Marber MS. New therapeutic targets in cardiology: p38 alpha mitogen-activated protein kinase for ischemic heart disease. Circulation. 2012;126:357–368. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
