

doi: 10.1038/embor.2013.106.
Epub 2013 Jul 23.
The ribosome triggers the stringent response by RelA via a highly distorted tRNA
Affiliations
- PMID: 23877429
- PMCID: PMC3790049
- DOI: 10.1038/embor.2013.106
Item in Clipboard
The ribosome triggers the stringent response by RelA via a highly distorted tRNA
EMBO Rep.
2013 Sep.
Abstract
The bacterial stringent response links nutrient starvation with the transcriptional control of genes. This process is initiated by the stringent factor RelA, which senses the presence of deacylated tRNA in the ribosome as a symptom of amino-acid starvation to synthesize the alarmone (p)ppGpp. Here we report a cryo-EM study of RelA bound to ribosomes bearing cognate, deacylated tRNA in the A-site. The data show that RelA on the ribosome stabilizes an unusual distorted form of the tRNA, with the acceptor arm making contact with RelA and far from its normal location in the peptidyl transferase centre.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

Biochemical characterization. (A) The normal elongation cycle of translation: a cognate, aminoacylated tRNA (purple) in complex with EF-Tu (orange) and GTP is delivered to the A-site of the ribosome (left). Following peptidyl transfer and the formation of hybrid tRNA states (middle), EF-G catalyses the translocation of tRNA and mRNA in the 30S subunit (right). (B) During starvation, an uncharged tRNA (purple) binds to the A-site of the ribosome in the absence of EF-Tu, leading to stalling. This stalled state is recognized by RelA (red). (C) Diagram representing the domain organization of the relA gene. The two constructs (NT-454-RelA and full length) used for this work are represented. (D) 4–12% SDS–PAGE loaded with the pull-downs of the full-length RelA (left) or with the truncated NT-454-RelA construct (right) with different programmed states of the 70 S. mRNA, messenger RNA; tRNA, transfer RNAs.
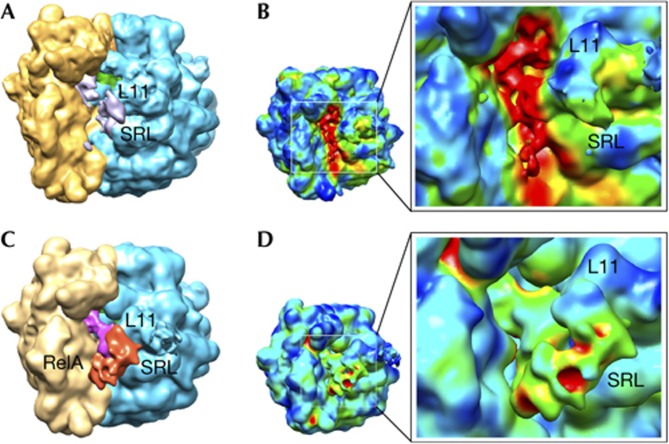
Cryo-EM reconstructions using the entire data set and after classification. (A) and (B) display renderings for the cryo-EM map calculated from the entire data set collected for the 70S–RelA complex. (A) Regions identified as the 30S subunit (yellow), 50S subunit (blue) and the P-site tRNA (green) are shown. Scattered signal attributable to the deacylated A-site tRNA and RelA is shown in grey. (B) 3D variance analysis showed that the regions of higher variability (shown in red) are located in the inter-subunit space. The reconstruction exhibits considerable heterogeneity, mostly located in the region of the A-site tRNA. Panels (C) and (D) show representations for the cryo-EM map with higher occupancy for RelA after classification of the original data set. (C) Orientation and labels as in panel (A) with additional densities attributable to the deacylated A-site tRNA (magenta) and protein RelA (red). (D) Conformational variability by 3D variance analysis suggested a clear improvement in the occupancy by RelA after the sorting. Labels indicate: L11 (region that contains ribosomal protein L11 and a segment of 23S rRNA); SRL (sarcin–ricin loop); and RelA, density attributed to the stringent factor RelA. rRNA, ribosomal RNA; SRL, sarcin–ricin loop; tRNA, transfer RNA.
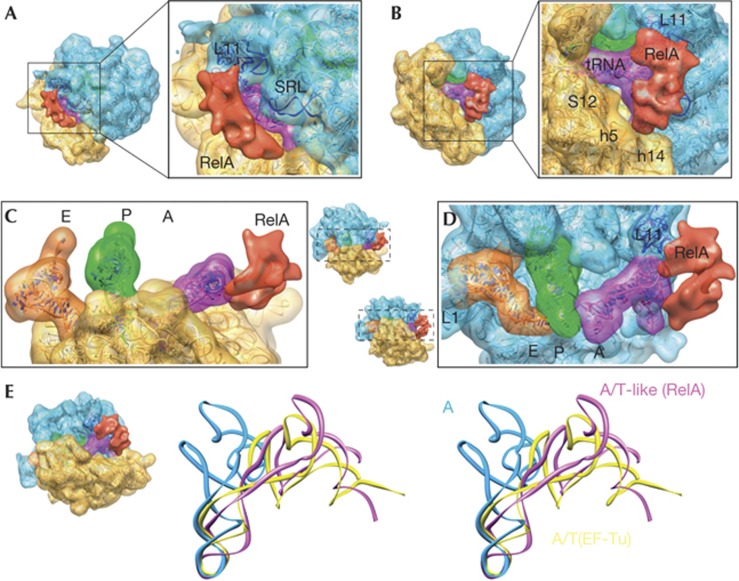
Three-dimensional cryo-EM structure of the 70S–RelA complex. (A–D) Cryo-EM reconstruction showing the regions corresponding to the 30S subunit (yellow), the 50S subunit (blue) and tRNAs in the A (magenta), P (green) and E (orange) sites. RelA is shown in red. The molecular structures fitted to the density are shown as ribbon diagrams. The isolated densities for the subunits, tRNAs and RelA are rendered using the same density threshold value. (E) Stereo views of the A-site tRNA in the current structure (magenta, denoted as A/T-like), the A/T state of aminoacyl–tRNA in complex with EF-Tu in the ribosome (yellow) [21] and the ‘classical’ A-site tRNA (blue) [38]. In each panel, the orientations of the subunits are shown as thumbnails. The following features are also labelled: L1 (stalk that contains ribosomal protein L1); SRL (sarcin–ricin loop); L11 (region that contains ribosomal protein L11 and a segment of 23S rRNA); S12 (ribosomal protein S12); h5 and h14 (helices from 16S rRNA). rRNA, ribosomal RNA; SRL, sarcin–ricin loop; tRNA, transfer RNAs
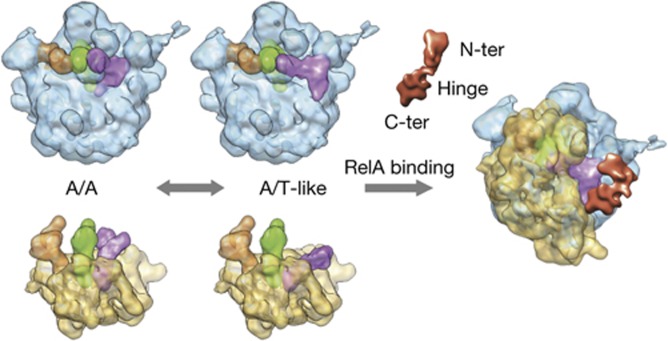
Cartoon-like schematic representation of RelA activation. See the text for details on the proposed model for the activation of RelA in binding to stalled ribosomes bearing deacylated tRNA.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
