

Six-rowed spike4 (Vrs4) controls spikelet determinacy and row-type in barley
- PMID: 23878219
- PMCID: PMC3740847
- DOI: 10.1073/pnas.1221950110
Six-rowed spike4 (Vrs4) controls spikelet determinacy and row-type in barley
Abstract
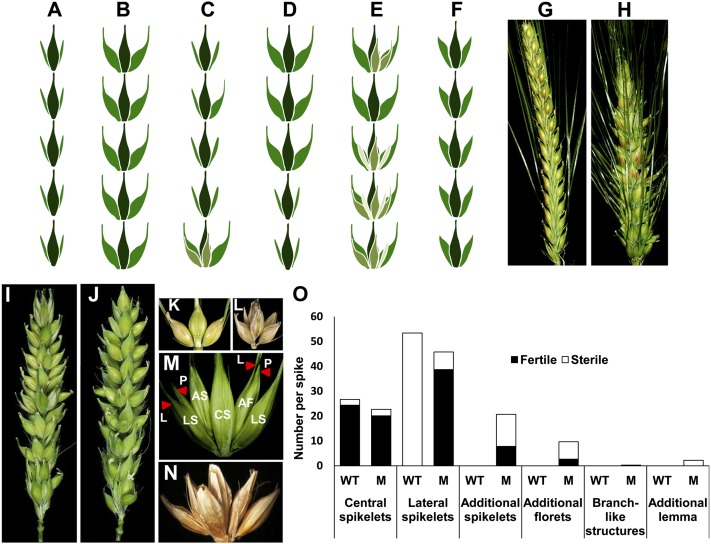
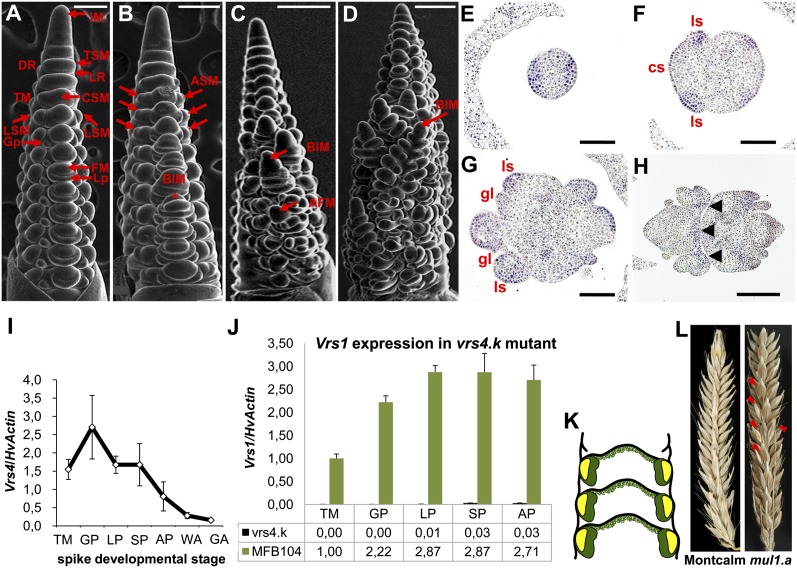
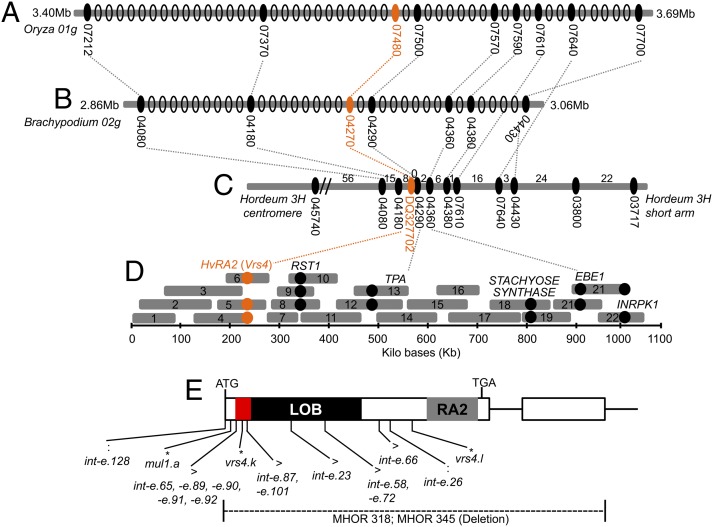
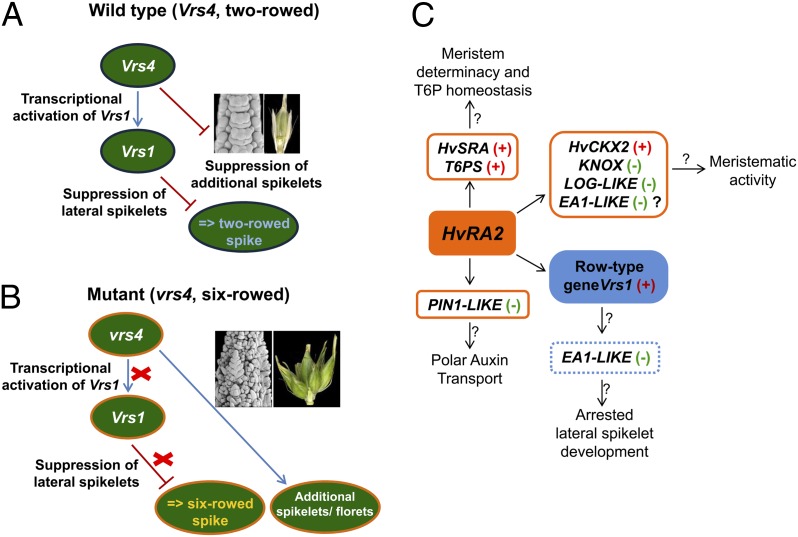
Inflorescence architecture of barley (Hordeum vulgare L.) is common among the Triticeae species, which bear one to three single-flowered spikelets at each rachis internode. Triple spikelet meristem is one of the unique features of barley spikes, in which three spikelets (one central and two lateral spikelets) are produced at each rachis internode. Fertility of the lateral spikelets at triple spikelet meristem gives row-type identity to barley spikes. Six-rowed spikes show fertile lateral spikelets and produce increased grain yield per spike, compared with two-rowed spikes with sterile lateral spikelets. Thus, far, two loci governing the row-type phenotype were isolated in barley that include Six-rowed spike1 (Vrs1) and Intermedium-C. In the present study, we isolated Six-rowed spike4 (Vrs4), a barley ortholog of the maize (Zea mays L.) inflorescence architecture gene RAMOSA2 (RA2). Eighteen coding mutations in barley RA2 (HvRA2) were specifically associated with lateral spikelet fertility and loss of spikelet determinacy. Expression analyses through mRNA in situ hybridization and microarray showed that Vrs4 (HvRA2) controls the row-type pathway through Vrs1 (HvHox1), a negative regulator of lateral spikelet fertility in barley. Moreover, Vrs4 may also regulate transcripts of barley SISTER OF RAMOSA3 (HvSRA), a putative trehalose-6-phosphate phosphatase involved in trehalose-6-phosphate homeostasis implicated to control spikelet determinacy. Our expression data illustrated that, although RA2 is conserved among different grass species, its down-stream target genes appear to be modified in barley and possibly other species of tribe Triticeae.
Keywords: EGG APPARATUS1; cytokinin; grain number; yield potential.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kellogg EA. Floral displays: Genetic control of grass inflorescences. Curr Opin Plant Biol. 2007;10(1):26–31. - PubMed
-
- Vollbrecht E, Springer PS, Goh L, Buckler ES, Martienssen R. Architecture of floral branch systems in maize and related grasses. Nature. 2005;436(7054):1119–1126. - PubMed
-
- Satoh-Nagasawa N, Nagasawa N, Malcomber S, Sakai H, Jackson D. A trehalose metabolic enzyme controls inflorescence architecture in maize. Nature. 2006;441(7090):227–230. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
