

Structural and mechanistic insights into VEGF receptor 3 ligand binding and activation
- PMID: 23878260
- PMCID: PMC3740881
- DOI: 10.1073/pnas.1301415110
Structural and mechanistic insights into VEGF receptor 3 ligand binding and activation
Abstract
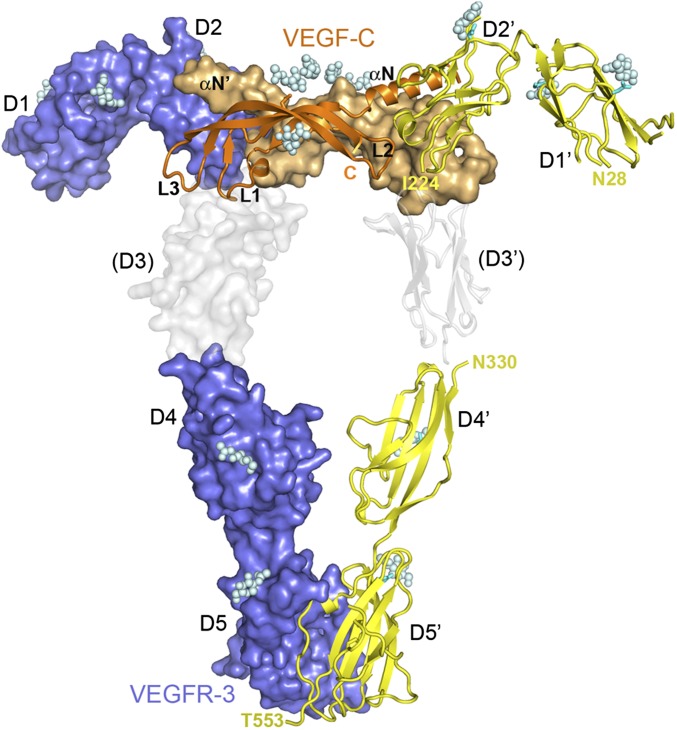
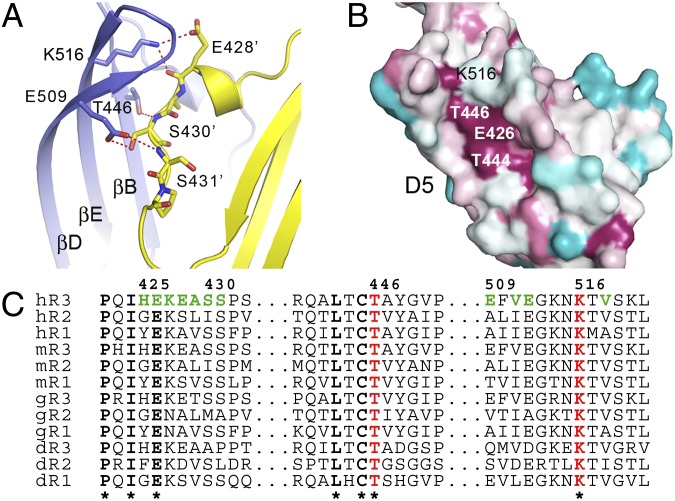
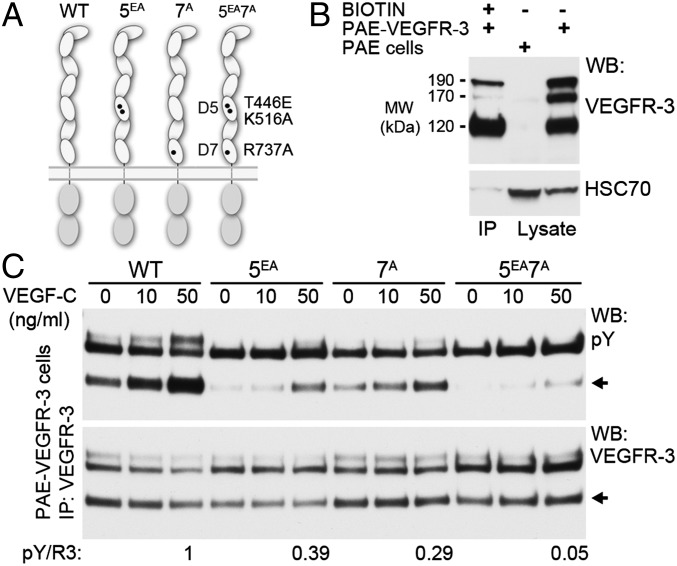
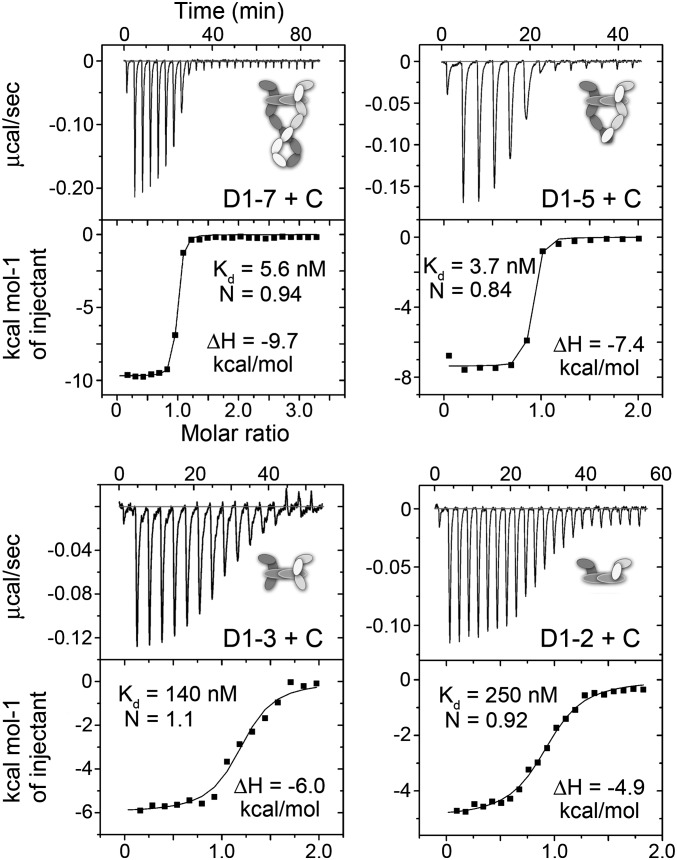
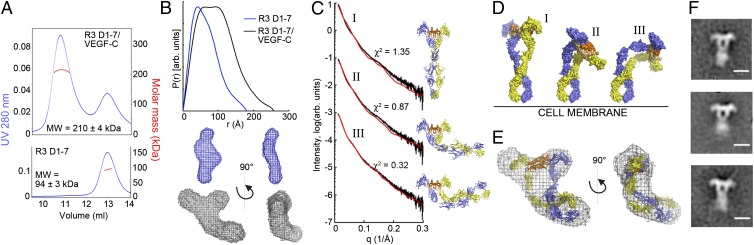
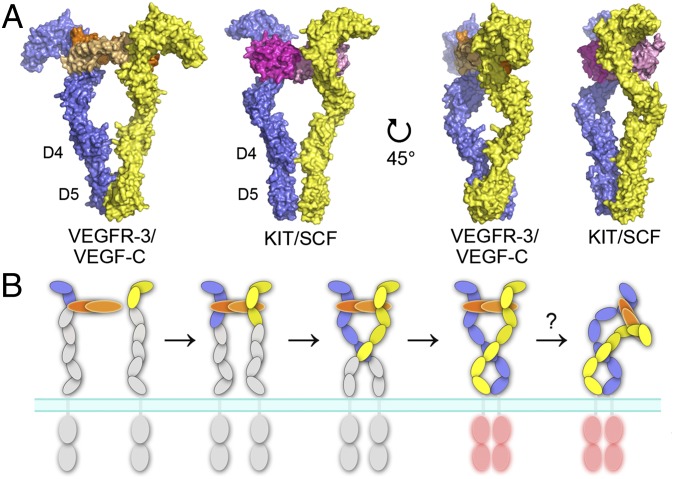
Vascular endothelial growth factors (VEGFs) and their receptors (VEGFRs) are key drivers of blood and lymph vessel formation in development, but also in several pathological processes. VEGF-C signaling through VEGFR-3 promotes lymphangiogenesis, which is a clinically relevant target for treating lymphatic insufficiency and for blocking tumor angiogenesis and metastasis. The extracellular domain of VEGFRs consists of seven Ig homology domains; domains 1-3 (D1-3) are responsible for ligand binding, and the membrane-proximal domains 4-7 (D4-7) are involved in structural rearrangements essential for receptor dimerization and activation. Here we analyzed the crystal structures of VEGF-C in complex with VEGFR-3 domains D1-2 and of the VEGFR-3 D4-5 homodimer. The structures revealed a conserved ligand-binding interface in D2 and a unique mechanism for VEGFR dimerization and activation, with homotypic interactions in D5. Mutation of the conserved residues mediating the D5 interaction (Thr446 and Lys516) and the D7 interaction (Arg737) compromised VEGF-C induced VEGFR-3 activation. A thermodynamic analysis of VEGFR-3 deletion mutants showed that D3, D4-5, and D6-7 all contribute to ligand binding. A structural model of the VEGF-C/VEGFR-3 D1-7 complex derived from small-angle X-ray scattering data is consistent with the homotypic interactions in D5 and D7. Taken together, our data show that ligand-dependent homotypic interactions in D5 and D7 are essential for VEGFR activation, opening promising possibilities for the design of VEGFR-specific drugs.
Keywords: receptor tyrosine kinase; signal transduction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structural determinants of growth factor binding and specificity by VEGF receptor 2.Proc Natl Acad Sci U S A. 2010 Feb 9;107(6):2425-30. doi: 10.1073/pnas.0914318107. Proc Natl Acad Sci U S A. 2010. PMID: 20145116 Free PMC article.
-
Thermodynamic and structural description of allosterically regulated VEGFR-2 dimerization.Blood. 2012 Feb 16;119(7):1781-8. doi: 10.1182/blood-2011-11-390922. Epub 2011 Dec 29. Blood. 2012. PMID: 22207738
-
Structural determinants of vascular endothelial growth factor-D receptor binding and specificity.Blood. 2011 Feb 3;117(5):1507-15. doi: 10.1182/blood-2010-08-301549. Epub 2010 Dec 8. Blood. 2011. PMID: 21148085
-
Targeting extracellular domains D4 and D7 of vascular endothelial growth factor receptor 2 reveals allosteric receptor regulatory sites.Mol Cell Biol. 2012 Oct;32(19):3802-13. doi: 10.1128/MCB.06787-11. Epub 2012 Jul 16. Mol Cell Biol. 2012. PMID: 22801374 Free PMC article. Review.
-
The role of the VEGF-C/VEGFR-3 axis in cancer progression.Br J Cancer. 2007 Feb 26;96(4):541-5. doi: 10.1038/sj.bjc.6603487. Epub 2006 Dec 12. Br J Cancer. 2007. PMID: 17164762 Free PMC article. Review.
Cited by
-
Regulation of oligomeric organization of the serotonin 5-hydroxytryptamine 2C (5-HT2C) receptor observed by spatial intensity distribution analysis.J Biol Chem. 2015 May 15;290(20):12844-57. doi: 10.1074/jbc.M115.644724. Epub 2015 Mar 30. J Biol Chem. 2015. PMID: 25825490 Free PMC article.
-
Plump endothelial cells integrated into pre-existing venules contribute to the formation of 'mother' and 'daughter' vessels in pyogenic granuloma: possible role of galectin-1, -3 and -8.Scars Burn Heal. 2021 Jan 22;7:2059513120986687. doi: 10.1177/2059513120986687. eCollection 2021 Jan-Dec. Scars Burn Heal. 2021. PMID: 33796337 Free PMC article.
-
Discovery of pan-VEGF inhibitory peptides directed to the extracellular ligand-binding domains of the VEGF receptors.Sci Adv. 2016 Oct 28;2(10):e1600611. doi: 10.1126/sciadv.1600611. eCollection 2016 Oct. Sci Adv. 2016. PMID: 27819042 Free PMC article.
-
Allosteric targeting of receptor tyrosine kinases.Nat Biotechnol. 2014 Nov;32(11):1113-20. doi: 10.1038/nbt.3028. Nat Biotechnol. 2014. PMID: 25380447
-
Biomacromolecules as carriers in drug delivery and tissue engineering.Acta Pharm Sin B. 2018 Jan;8(1):34-50. doi: 10.1016/j.apsb.2017.11.005. Epub 2017 Dec 9. Acta Pharm Sin B. 2018. PMID: 29872621 Free PMC article. Review.
References
-
- Koch S, Tugues S, Li X, Gualandi L, Claesson-Welsh L. Signal transduction by vascular endothelial growth factor receptors. Biochem J. 2011;437(2):169–183. - PubMed
-
- Ferrara N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr Rev. 2004;25(4):581–611. - PubMed
-
- Tammela T, Alitalo K. Lymphangiogenesis: Molecular mechanisms and future promise. Cell. 2010;140(4):460–476. - PubMed
-
- Ferrara N, Kerbel RS. Angiogenesis as a therapeutic target. Nature. 2005;438(7070):967–974. - PubMed
-
- Alitalo K. The lymphatic vasculature in disease. Nat Med. 2011;17(11):1371–1380. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
