

Allergens as immunomodulatory proteins: the cat dander protein Fel d 1 enhances TLR activation by lipid ligands
- PMID: 23878318
- PMCID: PMC3836235
- DOI: 10.4049/jimmunol.1300284
Allergens as immunomodulatory proteins: the cat dander protein Fel d 1 enhances TLR activation by lipid ligands
Abstract
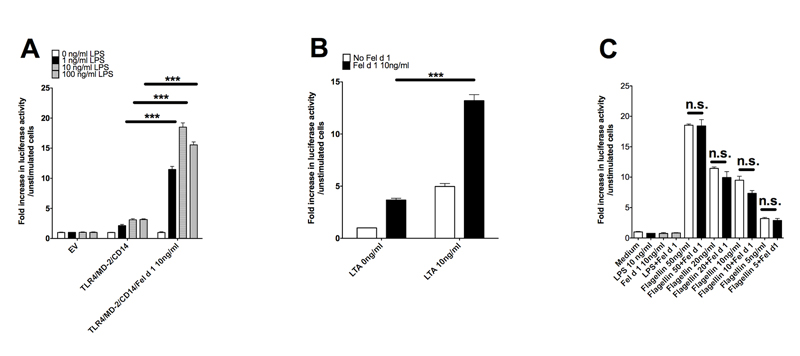
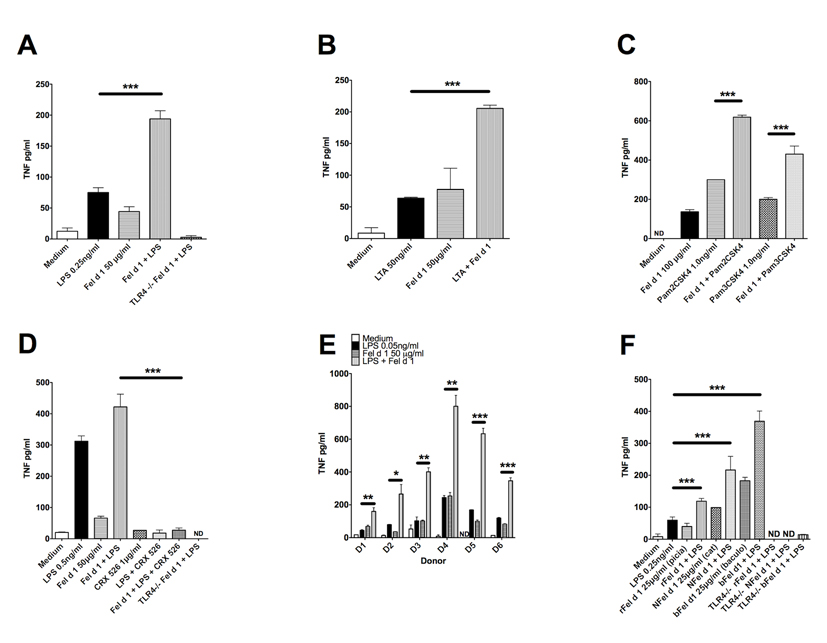
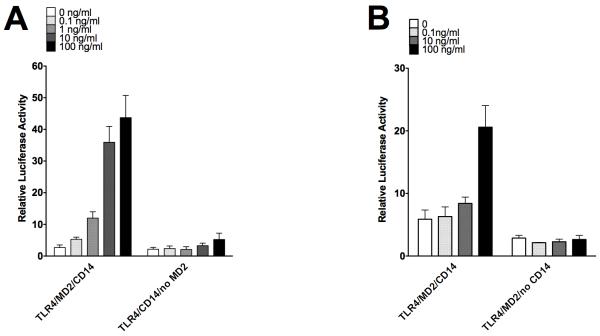
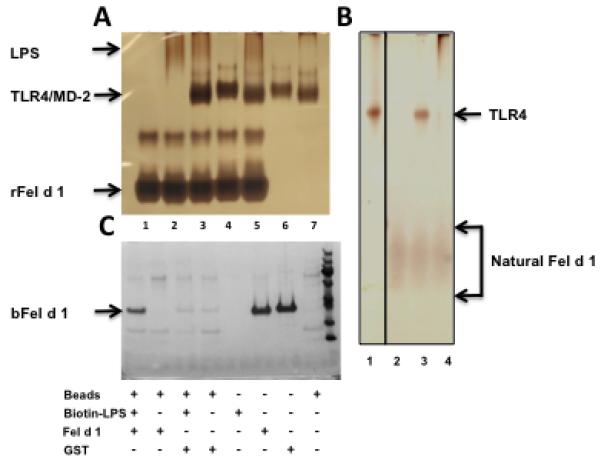
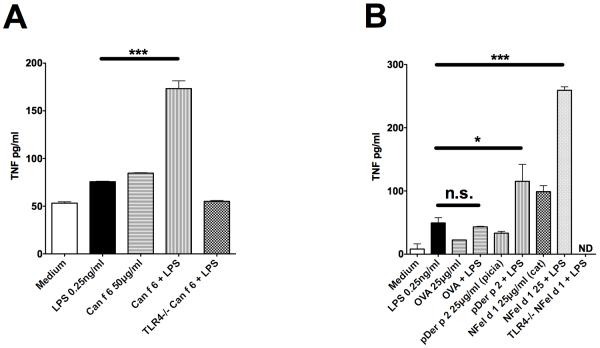
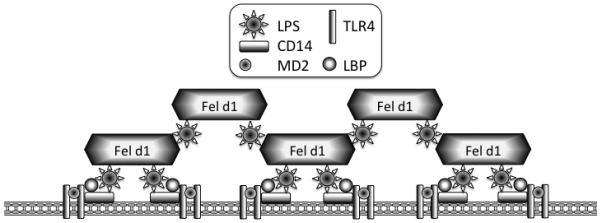
Allergic responses can be triggered by structurally diverse allergens. Most allergens are proteins, yet extensive research has not revealed how they initiate the allergic response and why the myriad of other inhaled proteins do not. Among these allergens, the cat secretoglobulin protein Fel d 1 is a major allergen and is responsible for severe allergic responses. In this study, we show that similar to the mite dust allergen Der p 2, Fel d 1 substantially enhances signaling through the innate receptors TLR4 and TLR2. In contrast to Der p 2, however, Fel d 1 does not act by mimicking the TLR4 coreceptor MD2 and is not able to bind stably to the TLR4/MD2 complex in vitro. Fel d 1 does, however, bind to the TLR4 agonist LPS, suggesting that a lipid transfer mechanism may be involved in the Fel d 1 enhancement of TLR signaling. We also show that the dog allergen Can f 6, a member of a distinct class of lipocalin allergens, has very similar properties to Fel d 1. We propose that Fel d 1 and Can f 6 belong to a group of allergen immunomodulatory proteins that enhance innate immune signaling and promote airway hypersensitivity reactions in diseases such as asthma.
Figures
References
-
- Minnicozzi M, Sawyer RT, Fenton MJ. Innate immunity in allergic disease. Immunological reviews. 2011;242:106–127. - PubMed
-
- Spitzauer S. Allergy to mammalian proteins: at the borderline between foreign and self? International archives of allergy and immunology. 1999;120:259–269. - PubMed
-
- Chiou Y-L, Lin C-Y. Der p2 activates airway smooth muscle cells in a TLR2/MyD88-dependent manner to induce an inflammatory response. Journal of cellular physiology. 2009;220:311–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
