

Contraction-induced lipolysis is not impaired by inhibition of hormone-sensitive lipase in skeletal muscle
- PMID: 23878361
- PMCID: PMC3810815
- DOI: 10.1113/jphysiol.2013.260794
Contraction-induced lipolysis is not impaired by inhibition of hormone-sensitive lipase in skeletal muscle
Abstract
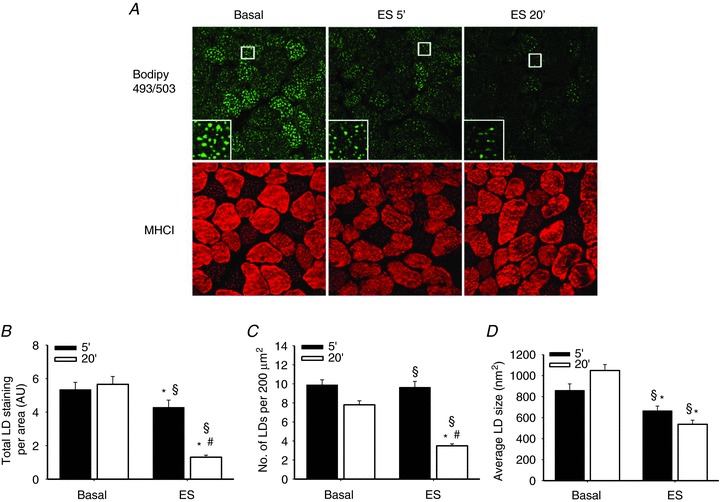
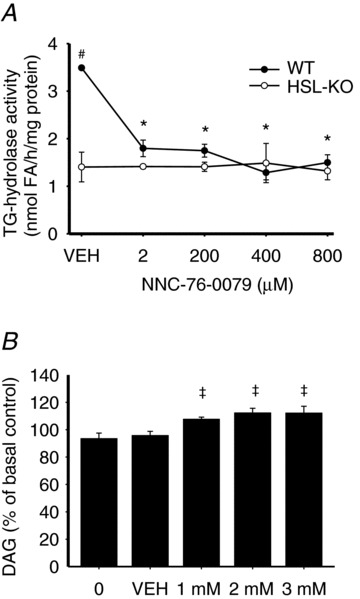
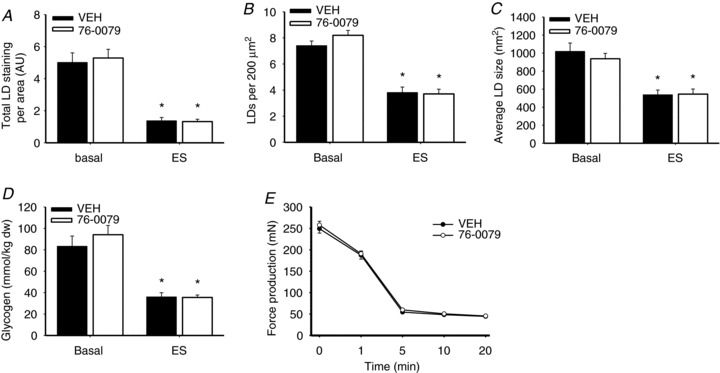
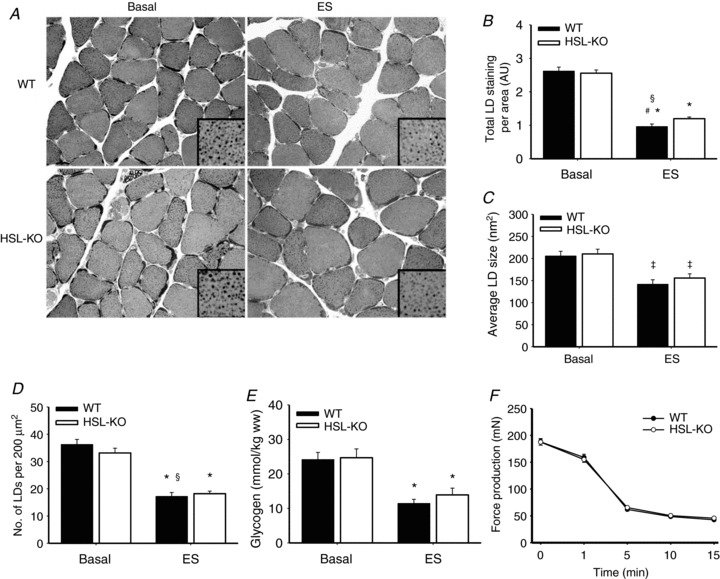
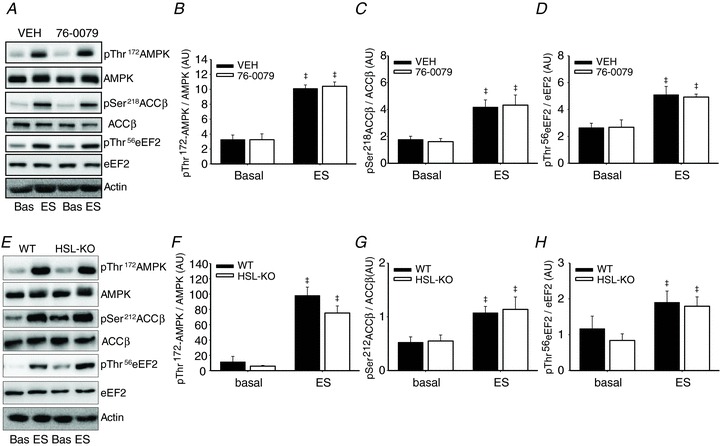
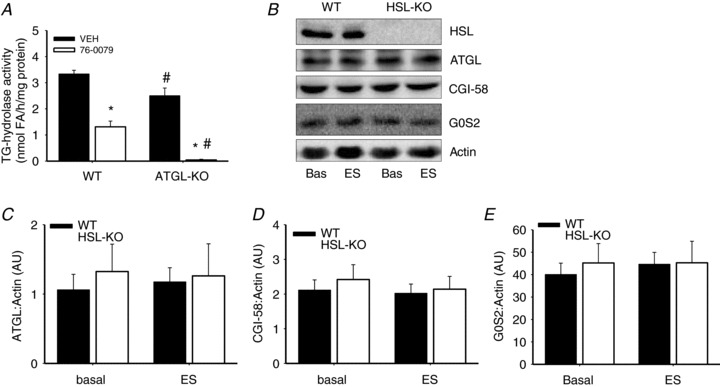
In skeletal muscle hormone-sensitive lipase (HSL) has long been accepted to be the principal enzyme responsible for lipolysis of intramyocellular triacylglycerol (IMTG) during contractions. However, this notion is based on in vitro lipase activity data, which may not reflect the in vivo lipolytic activity. We investigated lipolysis of IMTG in soleus muscles electrically stimulated to contract ex vivo during acute pharmacological inhibition of HSL in rat muscles and in muscles from HSL knockout (HSL-KO) mice. Measurements of IMTG are complicated by the presence of adipocytes located between the muscle fibres. To circumvent the problem with this contamination we analysed intramyocellular lipid droplet content histochemically. At maximal inhibition of HSL in rat muscles, contraction-induced breakdown of IMTG was identical to that seen in control muscles (P < 0.001). In response to contractions IMTG staining decreased significantly in both HSL-KO and WT muscles (P < 0.05). In vitro TG hydrolase activity data revealed that adipose triglyceride lipase (ATGL) and HSL collectively account for ∼98% of the TG hydrolase activity in mouse skeletal muscle, other TG lipases accordingly being of negligible importance for lipolysis of IMTG. The present study is the first to demonstrate that contraction-induced lipolysis of IMTG occurs in the absence of HSL activity in rat and mouse skeletal muscle. Furthermore, the results suggest that ATGL is activated and plays a major role in lipolysis of IMTG during muscle contractions.
Figures
Comment in
-
Lipid metabolism in contracting muscle--HSL takes a back seat.J Physiol. 2013 Oct 15;591(20):4951. doi: 10.1113/jphysiol.2013.262766. J Physiol. 2013. PMID: 24130317 Free PMC article. No abstract available.
-
HSL and ATGL: the movers and shakers of muscle lipolysis.J Physiol. 2013 Dec 15;591(24):6137-8. doi: 10.1113/jphysiol.2013.265199. J Physiol. 2013. PMID: 24339152 Free PMC article. No abstract available.
References
-
- Alsted TJ, Nybo L, Schweiger M, Fledelius C, Jacobsen P, Zimmermann R, Zechner R, Kiens B. Adipose triglyceride lipase in human skeletal muscle is upregulated by exercise training. Am J Physiol Endocrinol Metab. 2009;296:E445–453. - PubMed
-
- Bergstrom J, Hultman E, Saltin B. Muscle glycogen consumption during cross-country skiing (the Vasa ski race) Int Z Angew Physiol. 1973;31:71–75. - PubMed
-
- Bezaire V, Mairal A, Ribet C, Lefort C, Girousse A, Jocken J, Laurencikiene J, Anesia R, Rodriguez AM, Ryden M, Stenson BM, Dani C, Ailhaud G, Arner P, Langin D. Contribution of adipose triglyceride lipase and hormone-sensitive lipase to lipolysis in hMADS adipocytes. J Biol Chem. 2009;284:18282–18291. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous