

Biogenesis, turnover, and mode of action of plant microRNAs
- PMID: 23881412
- PMCID: PMC3753372
- DOI: 10.1105/tpc.113.113159
Biogenesis, turnover, and mode of action of plant microRNAs
Abstract
MicroRNAs (miRNAs) are small RNAs that control gene expression through silencing of target mRNAs. Mature miRNAs are processed from primary miRNA transcripts by the endonuclease activity of the DICER-LIKE1 (DCL1) protein complex. Mechanisms exist that allow the DCL1 complex to precisely excise the miRNA from its precursor. Our understanding of miRNA biogenesis, particularly its intersection with transcription and other aspects of RNA metabolism such as splicing, is still evolving. Mature miRNAs are incorporated into an ARGONAUTE (AGO) effector complex competent for target gene silencing but are also subjected to turnover through a degradation mechanism that is beginning to be understood. The mechanisms of miRNA target silencing in plants are no longer limited to AGO-catalyzed slicing, and the contribution of translational inhibition is increasingly appreciated. Here, we review the mechanisms underlying the biogenesis, turnover, and activities of plant miRNAs.
Figures
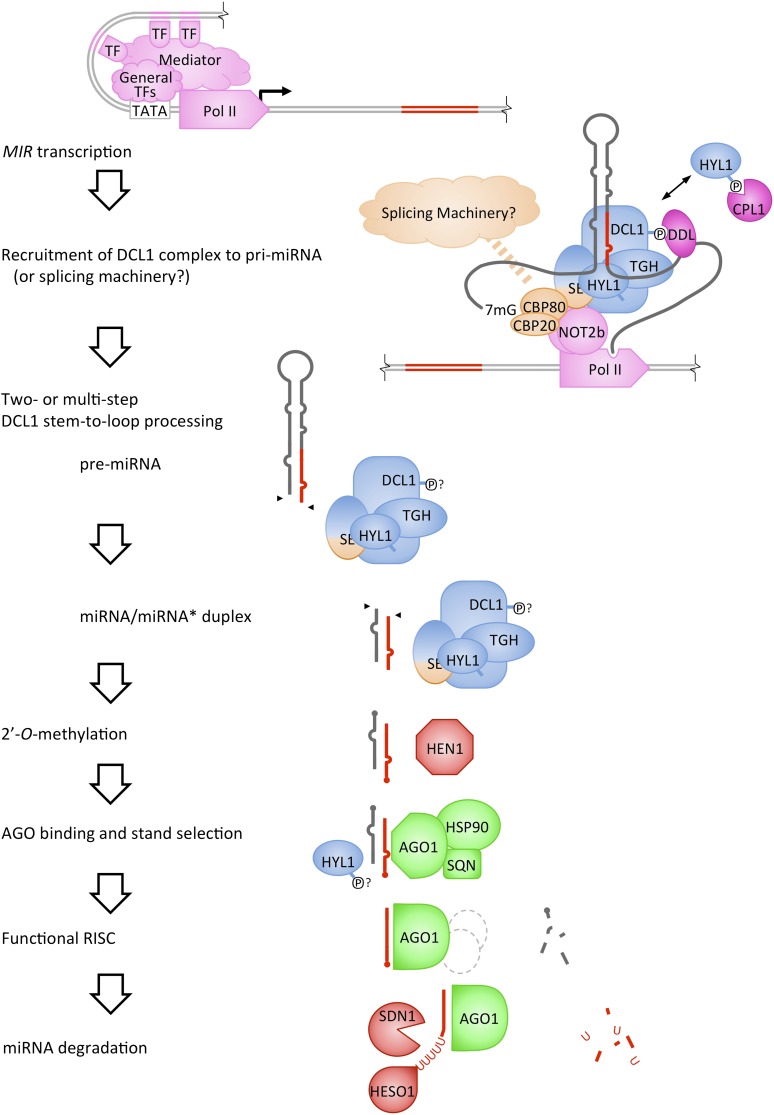
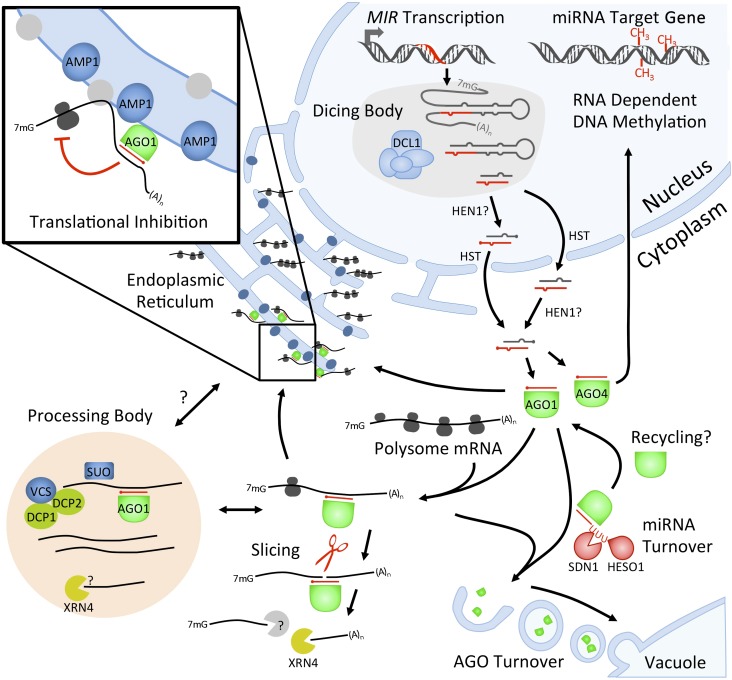
Comment in
-
The plant cell reviews aspects of microRNA and PhasiRNA regulatory function.Plant Cell. 2013 Jul;25(7):2382. doi: 10.1105/tpc.113.250713. Epub 2013 Jul 29. Plant Cell. 2013. PMID: 23897925 Free PMC article. No abstract available.
References
-
- Allen E., Xie Z., Gustafson A.M., Carrington J.C. (2005). MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
