

Review
doi: 10.1101/cshperspect.a012807.
The minichromosome maintenance replicative helicase
Affiliations
- PMID: 23881943
- PMCID: PMC3809582
- DOI: 10.1101/cshperspect.a012807
Item in Clipboard
Review
The minichromosome maintenance replicative helicase
Cold Spring Harb Perspect Biol.
.
Abstract
The eukaryotic replicative helicase, the minichromosome maintenance (MCM) complex, is composed of six distinct, but related, subunits MCM(2-7). The relationship between the sequences of the subunits indicates that they are derived from a common ancestor and indeed, present-day archaea possess a homohexameric MCM. Recent progress in the biochemical and structural studies of both eukaryal and archaeal MCM complexes are beginning to shed light on the mechanisms of action of this key component of the replisome.
Figures
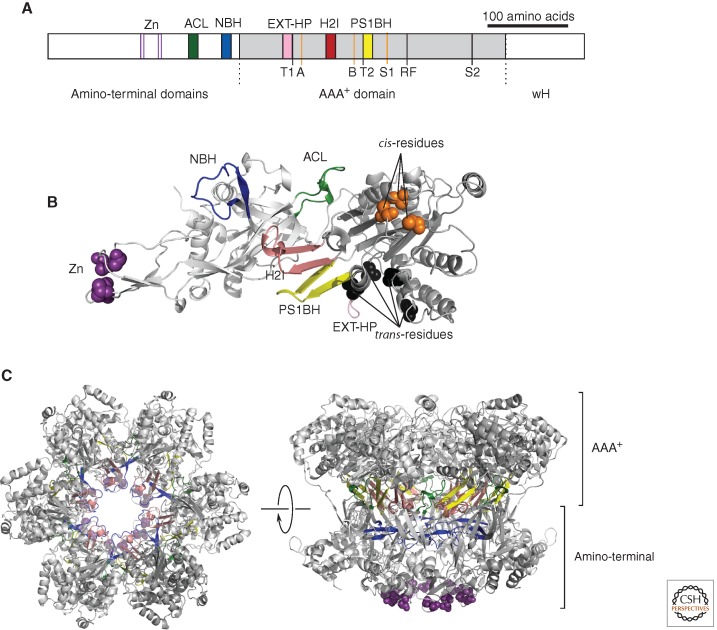
(A) Linear representation of a monomer of the archaeal MCM. (Gray) The central AAA+ domain; (white) the flanking amino-terminal domains and winged helix (wH). The position of key secondary structural elements—(Zn) zinc-binding; (ACL) allosteric communication loop; (NBH) amino-terminal β-hairpin; (EXT-HP) external β-hairpin; (H2I) helix 2 insert; and (PS1BH) pre-sensor 1 β-hairpin—are indicated above the figure and shown by colored blocks, the colors corresponding to those used in panels B and C. Key residues involved in the ATPase active site are indicated below the figure. (Orange lines) A, B, and S1, shown as orange lines are the Walker A lysine (K346), Walker B glutamate (E404), and Sensor 1 asparagine (N448), respectively, and constitute “cis”-acting residues. (Black lines) “Trans”-acting residues T1 (R331), T2 (Q423), arginine finger (R473), and Sensor 2 (R560). Numbering is from SsoMCM. (B) Structure of a monomer of SsoMCM (lacking detail of the wH domain). Secondary structure elements are labeled and colored in cartoon format, using the color scheme in panel A. (Purple spheres) The atoms of the zinc-coordinating residues; (orange spheres) the cis-acting residues; (black spheres) the trans-residues. (C) Model of a symmetric hexamer of SsoMCM. (Left) View down the central cavity of SsoMCM, looking from the carboxy-terminal face. (Right) The same hexamer rotated 90° to show a side view. The two tiers corresponding to the amino-terminal and AAA+ domains are indicated. The color scheme is as in panels A and B. Panels B and C were generated from PDB entry 3F9V using PyMOL (http://www.pymol.org ).
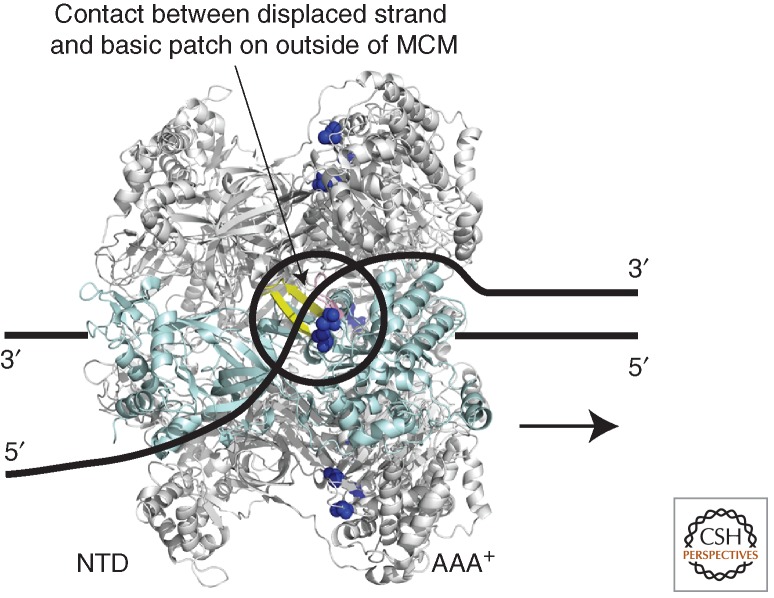
Model for MCM's helicase activity. MCM is shown to 3′-to-5′ translocate along a single strand of DNA that passes through the central cavity of the hexamer, one subunit of which is shown in pale blue. The displaced strand makes transient contacts with the outer surface of the hexamer (Rothenberg et al. 2007; Graham et al. 2011). Residues K323 and R440 of SsoMCM have been implicated in this contact (dark blue spheres) (Graham et al. 2012). (Yellow ribbon) The adjacent PS1BH. Aside from the potential role of these charged residues in contacting the displaced strand, nothing is known regarding the details of the trajectory of the displaced strand on the outer surface of the helicase.
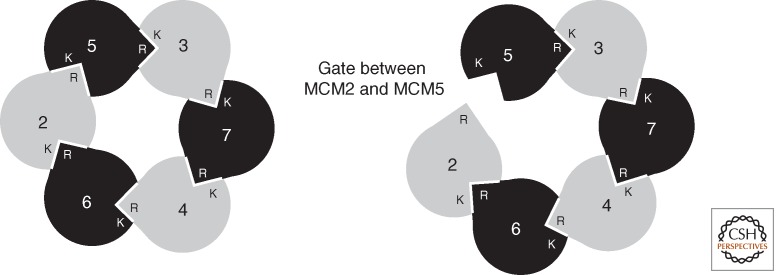
Organization of eukaryotic MCM(2–7). Interactions between protomers of MCM result in a heterohexamer of defined geometry (Ishimi 1997; Schwacha and Bell 2001; Davey et al. 2003). In the cartoon, the Ks indicate the cis-acting Walker A lysine residue of a given subunit, and the Rs show the trans-acting arginine fingers. Thus, the active site between protomers is contributed to by the cis surface of one protomer and the trans surface of its neighbor. As detailed in the text, considerable biochemical and structural data support the existence of a gate between subunits Mcm2 and Mcm5 (right panel).
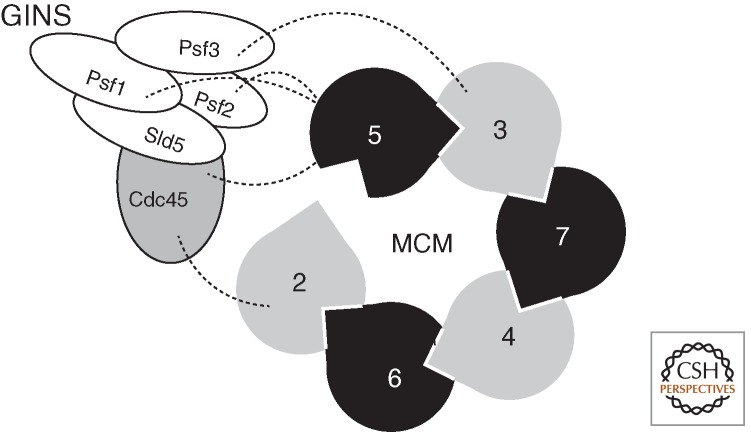
Interaction of the GINS components and Cdc45 with MCM. The cartoon indicates the open form of MCM(2–7) with dotted lines revealing the interactions between MCM subunits and Psf1, 2, 3, and Cdc45.
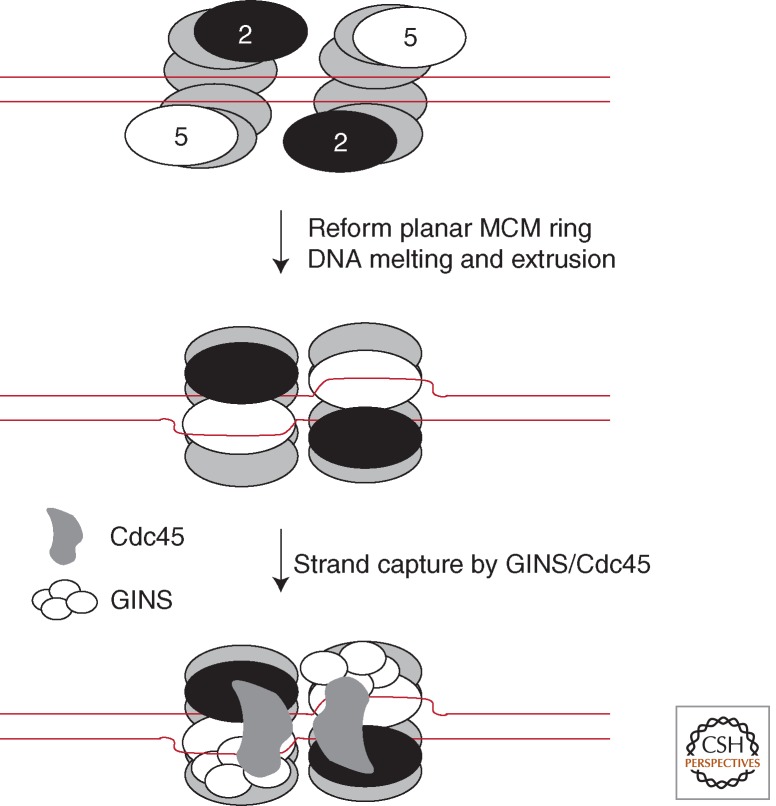
A speculative model for MCM loading and DNA melting at replication origins in eukarya and, by inference, archaea. Two open-gate hexamers of MCM are loaded at an origin. Ring closure occurs, concomitant with localized melting of DNA. One strand, the leading-strand template, is retained in the heart of the helicase; the other, the extruded lagging-strand template, is captured by the GINS/Cdc45 on the outer surface of the helicase assembly.
References
-
- Bae B, Chen Y-H, Costa A, Onesti S, Brunzelle JS, Lin Y, Cann IKO, Nair SK 2009. Insights into the architecture of the replicative helicase from the structure of an Archaeal MCM homolog. Structure 17: 211–222 - PubMed
-
- Beattie TR, Bell SD 2011. Molecular machines in Archaeal DNA replication. Curr Opin Chem Biol 15: 614–619 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases