

Genetic basis of cohesinopathies
- PMID: 23882154
- PMCID: PMC3711096
- DOI: 10.2147/TACG.S34457
Genetic basis of cohesinopathies
Abstract
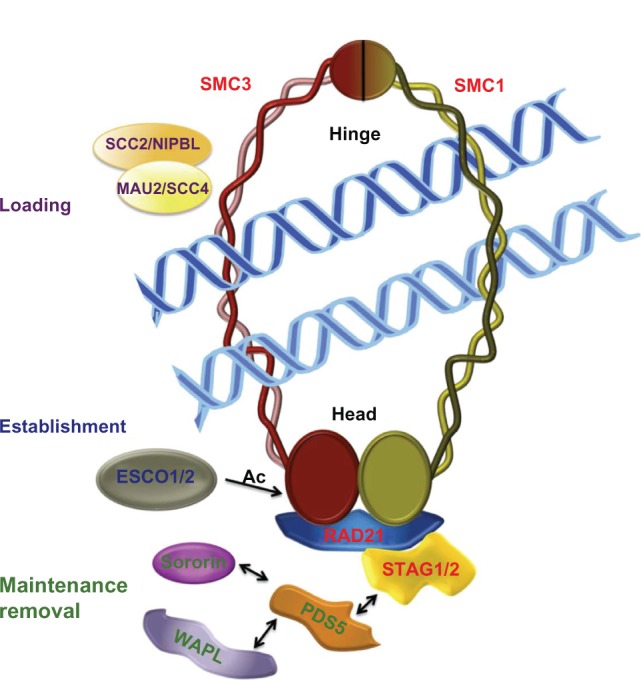
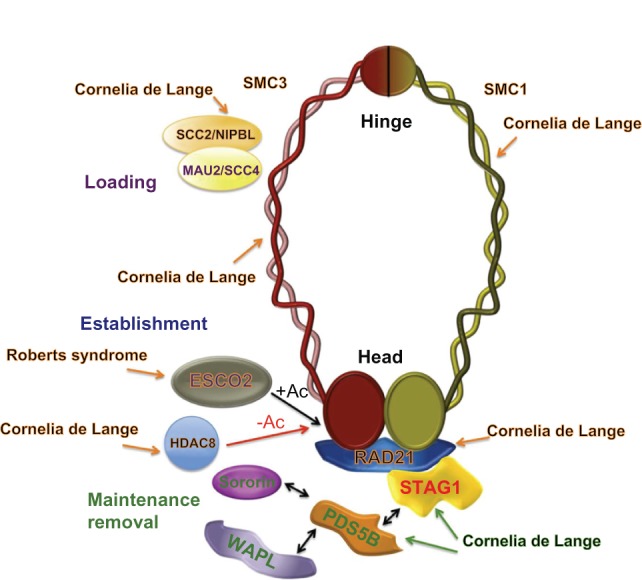
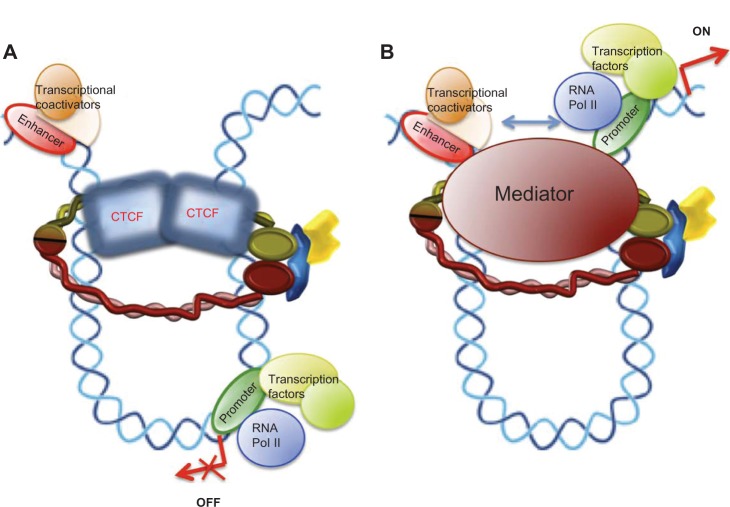
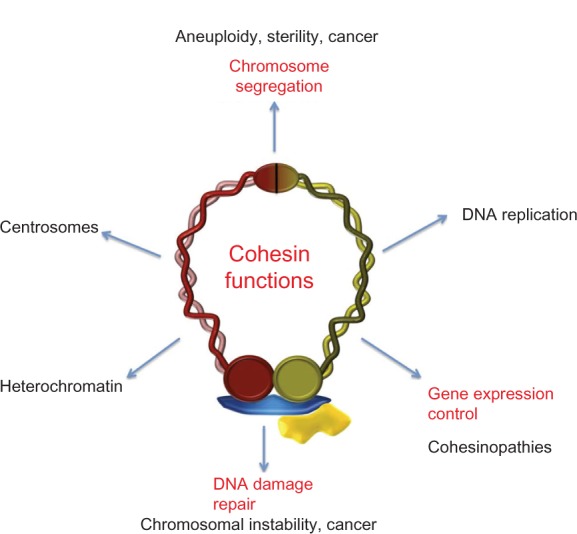
Cohesin is a ring-form multifunctional protein complex, which was discovered during a search for molecules that keep sister chromatids together during segregation of chromosomes during cell division. In the past decade, a large number of results have also demonstrated a need for the cohesin complex in other crucial events in the life cycle of the cell, including DNA duplication, heterochromatin formation, DNA double-strand break repair, and control of gene expression. The dynamics of the cohesin ring are modulated by a number of accessory and regulatory proteins, known as cohesin cofactors. Loss of function of the cohesin complex is incompatible with life; however, mutations in the genes encoding for cohesin subunits and/or cohesin cofactors, which have very little or a null effect on chromosome segregation, represent a newly recognized class of human genetic disorders known as cohesinopathies. A number of genetic, biochemical, and clinical approaches, and importantly, animal models, can help us to determine the underlying mechanisms for these human diseases.
Keywords: Cornelia de Lange syndrome; Roberts syndrome; cohesin; cohesinopathies; control; gene expression; insulators.
Figures
Similar articles
-
The expanding phenotypes of cohesinopathies: one ring to rule them all!Cell Cycle. 2019 Nov;18(21):2828-2848. doi: 10.1080/15384101.2019.1658476. Epub 2019 Sep 13. Cell Cycle. 2019. PMID: 31516082 Free PMC article. Review.
-
Cohesin: functions beyond sister chromatid cohesion.FEBS Lett. 2013 Aug 2;587(15):2299-312. doi: 10.1016/j.febslet.2013.06.035. Epub 2013 Jul 4. FEBS Lett. 2013. PMID: 23831059 Review.
-
Cohesinopathies: One ring, many obligations.Mutat Res. 2008 Dec 1;647(1-2):103-11. doi: 10.1016/j.mrfmmm.2008.08.010. Epub 2008 Aug 22. Mutat Res. 2008. PMID: 18786550 Review.
-
Roberts syndrome: A deficit in acetylated cohesin leads to nucleolar dysfunction.Rare Dis. 2014 Jan 21;2:e27743. doi: 10.4161/rdis.27743. eCollection 2014. Rare Dis. 2014. PMID: 25054091 Free PMC article.
-
Genome stability: What we have learned from cohesinopathies.Am J Med Genet C Semin Med Genet. 2016 Jun;172(2):171-8. doi: 10.1002/ajmg.c.31492. Epub 2016 Apr 19. Am J Med Genet C Semin Med Genet. 2016. PMID: 27091086 Review.
Cited by
-
Mutations in STAG2 cause an X-linked cohesinopathy associated with undergrowth, developmental delay, and dysmorphia: Expanding the phenotype in males.Mol Genet Genomic Med. 2019 Feb;7(2):e00501. doi: 10.1002/mgg3.501. Epub 2018 Nov 16. Mol Genet Genomic Med. 2019. PMID: 30447054 Free PMC article.
-
The Interplay of Cohesin and RNA Processing Factors: The Impact of Their Alterations on Genome Stability.Int J Mol Sci. 2022 Apr 1;23(7):3939. doi: 10.3390/ijms23073939. Int J Mol Sci. 2022. PMID: 35409298 Free PMC article. Review.
-
Positive Selection Drives the Evolution of the Structural Maintenance of Chromosomes (SMC) Complexes.Genes (Basel). 2024 Sep 3;15(9):1159. doi: 10.3390/genes15091159. Genes (Basel). 2024. PMID: 39336750 Free PMC article.
-
A novel STAG1 variant associated with congenital clubfoot and microphthalmia: A case report.SAGE Open Med Case Rep. 2024 Aug 31;12:2050313X241277123. doi: 10.1177/2050313X241277123. eCollection 2024. SAGE Open Med Case Rep. 2024. PMID: 39224759 Free PMC article.
-
The cohesin complex in mammalian meiosis.Genes Cells. 2019 Jan;24(1):6-30. doi: 10.1111/gtc.12652. Epub 2018 Nov 27. Genes Cells. 2019. PMID: 30479058 Free PMC article. Review.
References
-
- Haering CH, Jessberger R. Cohesin in determining chromosome architecture. Exp Cell Res. 2012;318:1386–1393. - PubMed
-
- Xiong B, Gerton JL. Regulators of cohesin network. Annu Rev Biochem. 2010;79:131–153. - PubMed
-
- Strom L, Lindroos HB, Shirahige K, Sjogren C. Postreplicative recruitment of cohesin to double-strand breaks is required for DNA repair. Mol Cell. 2004;16:1003–1015. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
