

Ganglioside GM1-mediated transcytosis of cholera toxin bypasses the retrograde pathway and depends on the structure of the ceramide domain
- PMID: 23884419
- PMCID: PMC3764787
- DOI: 10.1074/jbc.M113.474957
Ganglioside GM1-mediated transcytosis of cholera toxin bypasses the retrograde pathway and depends on the structure of the ceramide domain
Abstract
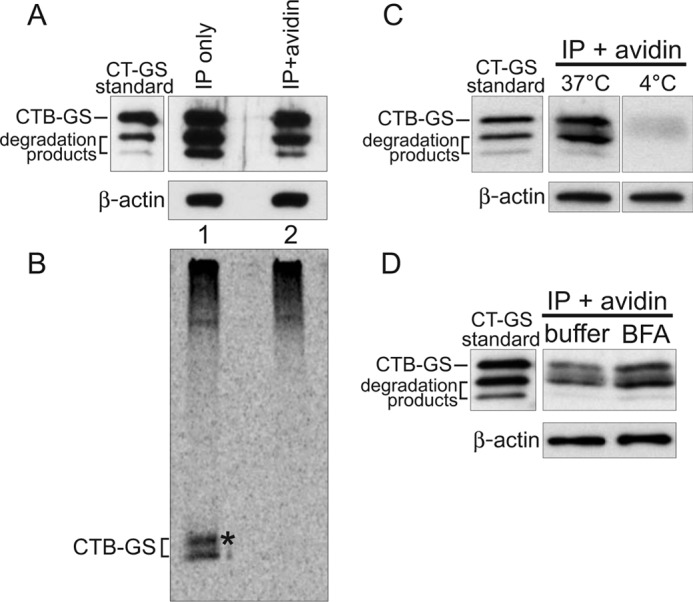
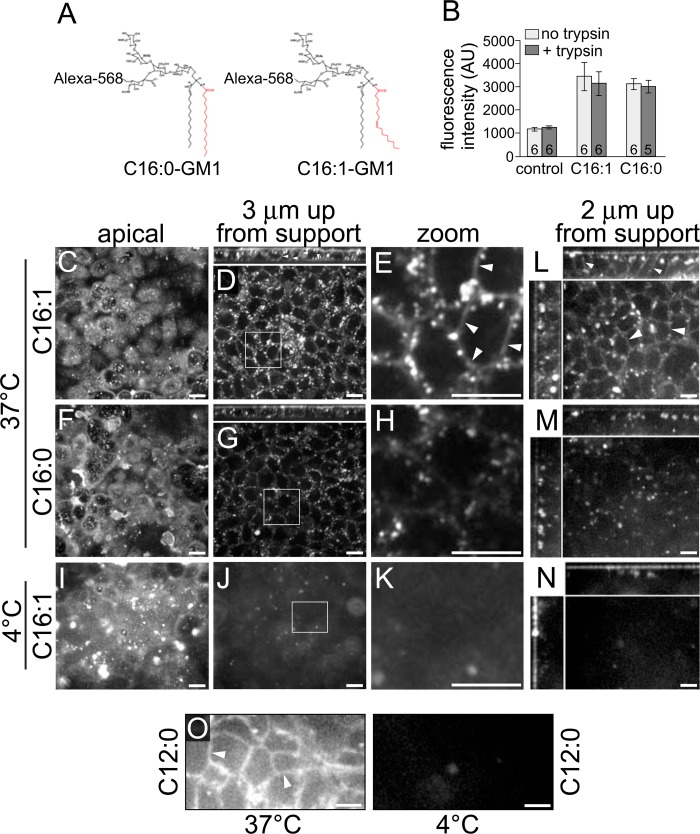
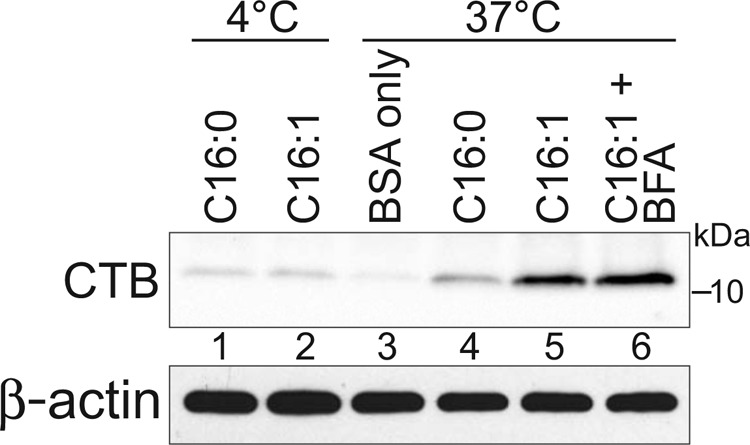
Cholera toxin causes diarrheal disease by binding ganglioside GM1 on the apical membrane of polarized intestinal epithelial cells and trafficking retrograde through sorting endosomes, the trans-Golgi network (TGN), and into the endoplasmic reticulum. A fraction of toxin also moves from endosomes across the cell to the basolateral plasma membrane by transcytosis, thus breeching the intestinal barrier. Here we find that sorting of cholera toxin into this transcytotic pathway bypasses retrograde transport to the TGN. We also find that GM1 sphingolipids can traffic from apical to basolateral membranes by transcytosis in the absence of toxin binding but only if the GM1 species contain cis-unsaturated or short acyl chains in the ceramide domain. We found previously that the same GM1 species are needed to efficiently traffic retrograde into the TGN and endoplasmic reticulum and into the recycling endosome, implicating a shared mechanism of action for sorting by lipid shape among these pathways.
Keywords: Cell Biology; Cholera Toxin; Endosomes; Ganglioside; Intracellular Trafficking; Lipid Transport; Membrane Lipids; Membrane Trafficking; Polarized Epithelia; Sphingolipid.
Figures
References
-
- Chinnapen D. J., Hsieh W. T., te Welscher Y. M., Saslowsky D. E., Kaoutzani L., Brandsma E., D'Auria L., Park H., Wagner J. S., Drake K. R., Kang M., Benjamin T., Ullman M. D., Costello C. E., Kenworthy A. K., Baumgart T., Massol R. H., Lencer W. I. (2012) Lipid sorting by ceramide structure from plasma membrane to ER for the cholera toxin receptor ganglioside GM1. Dev. Cell 23, 573–586 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
