

Crystal structure of 3WJ core revealing divalent ion-promoted thermostability and assembly of the Phi29 hexameric motor pRNA
- PMID: 23884902
- PMCID: PMC3753930
- DOI: 10.1261/rna.037077.112
Crystal structure of 3WJ core revealing divalent ion-promoted thermostability and assembly of the Phi29 hexameric motor pRNA
Abstract
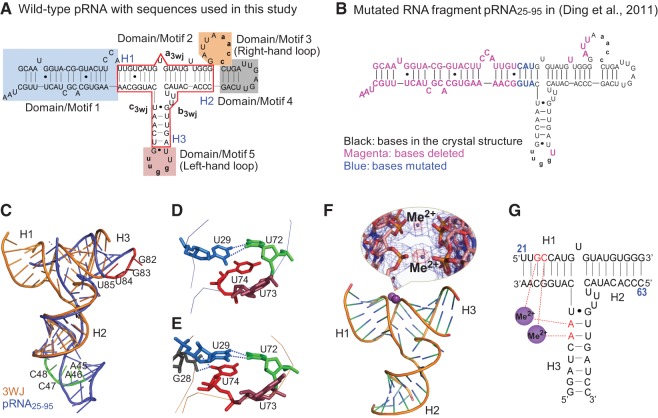
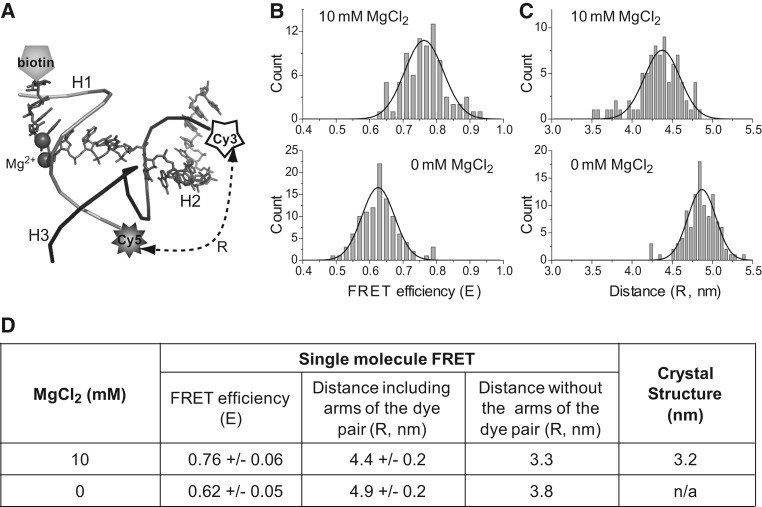
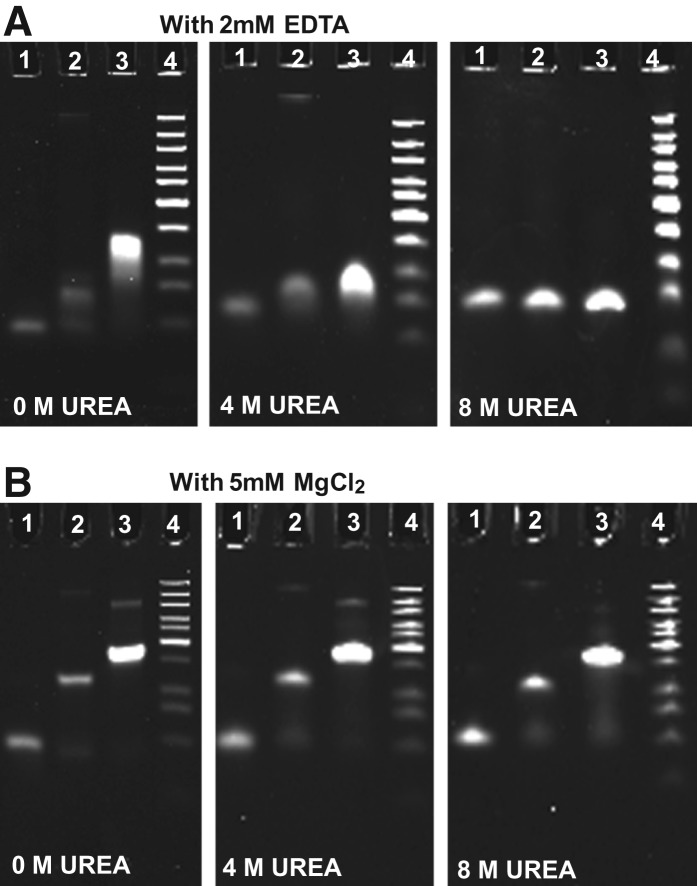
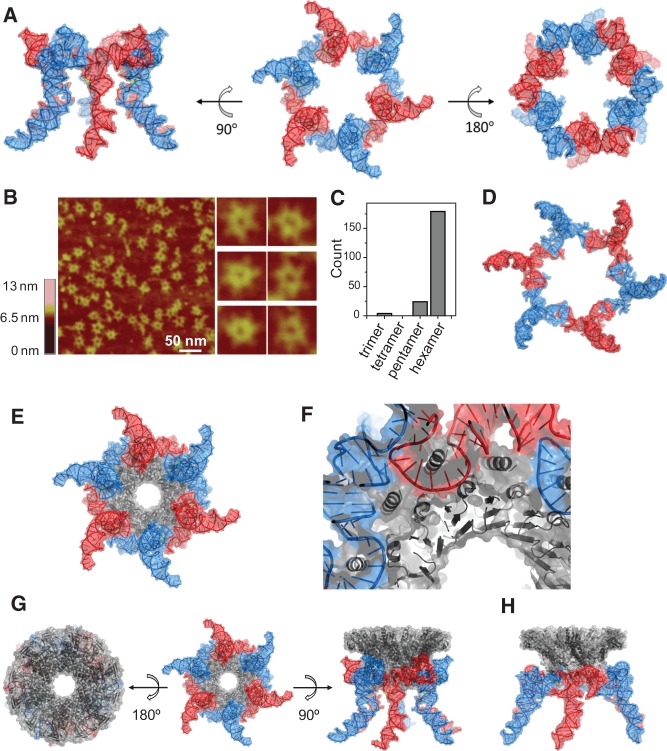
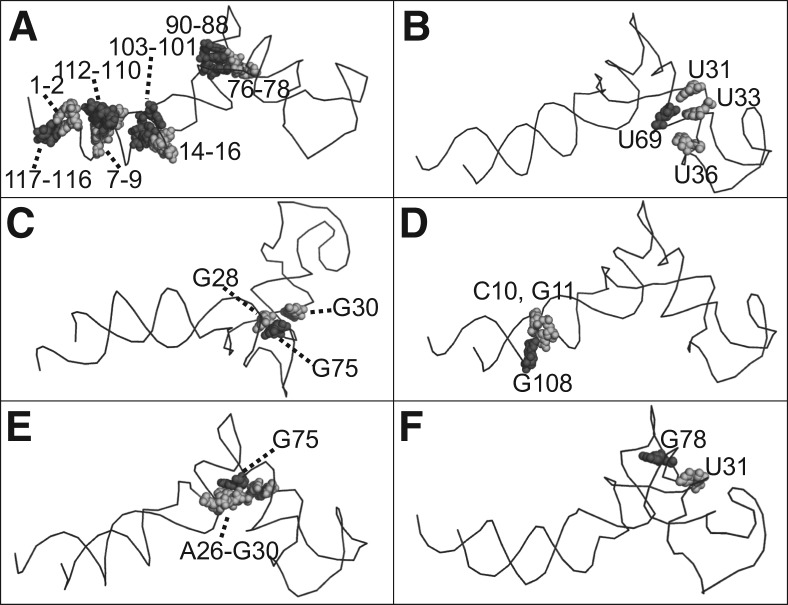
The bacteriophage phi29 DNA packaging motor, one of the strongest biological motors characterized to date, is geared by a packaging RNA (pRNA) ring. When assembled from three RNA fragments, its three-way junction (3WJ) motif is highly thermostable, is resistant to 8 M urea, and remains associated at extremely low concentrations in vitro and in vivo. To elucidate the structural basis for its unusual stability, we solved the crystal structure of this pRNA 3WJ motif at 3.05 Å. The structure revealed two divalent metal ions that coordinate 4 nt of the RNA fragments. Single-molecule fluorescence resonance energy transfer (smFRET) analysis confirmed a structural change of 3WJ upon addition of Mg²⁺. The reported pRNA 3WJ conformation is different from a previously published construct that lacks the metal coordination sites. The phi29 DNA packaging motor contains a dodecameric connector at the vertex of the procapsid, with a central pore for DNA translocation. This portal connector serves as the foothold for pRNA binding to procapsid. Subsequent modeling of a connector/pRNA complex suggests that the pRNA of the phi29 DNA packaging motor exists as a hexameric complex serving as a sheath over the connector. The model of hexameric pRNA on the connector agrees with AFM images of the phi29 pRNA hexamer acquired in air and matches all distance parameters obtained from cross-linking, complementary modification, and chemical modification interference.
Keywords: RNA crystal; RNA nanotechnology; RNA therapeutics; metal ions; three-way junction.
Figures
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC 2002. PHENIX: Building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr 58: 1948–1954 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous