

Type 1 inositol trisphosphate receptor regulates cerebellar circuits by maintaining the spine morphology of purkinje cells in adult mice
- PMID: 23884927
- PMCID: PMC6618669
- DOI: 10.1523/JNEUROSCI.0545-13.2013
Type 1 inositol trisphosphate receptor regulates cerebellar circuits by maintaining the spine morphology of purkinje cells in adult mice
Abstract
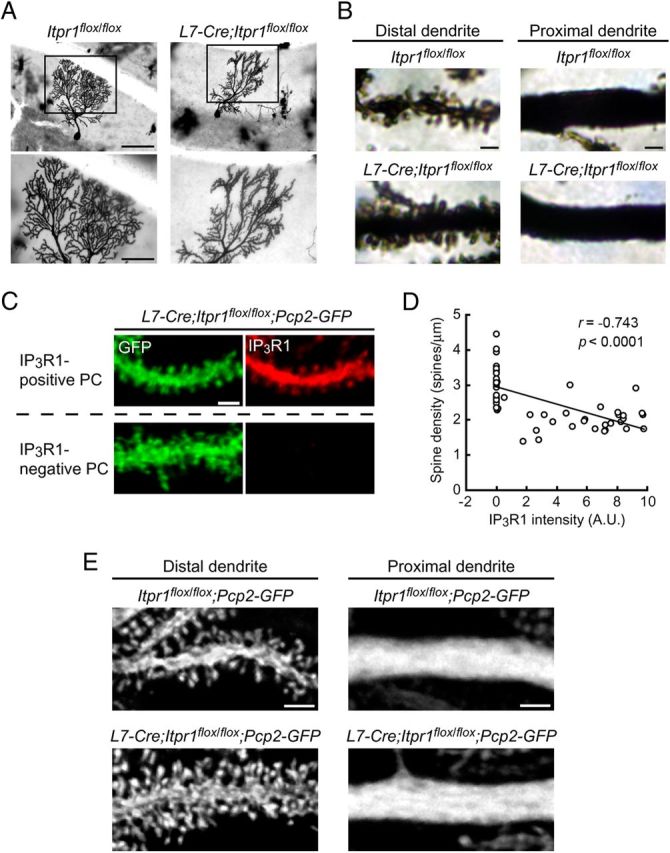
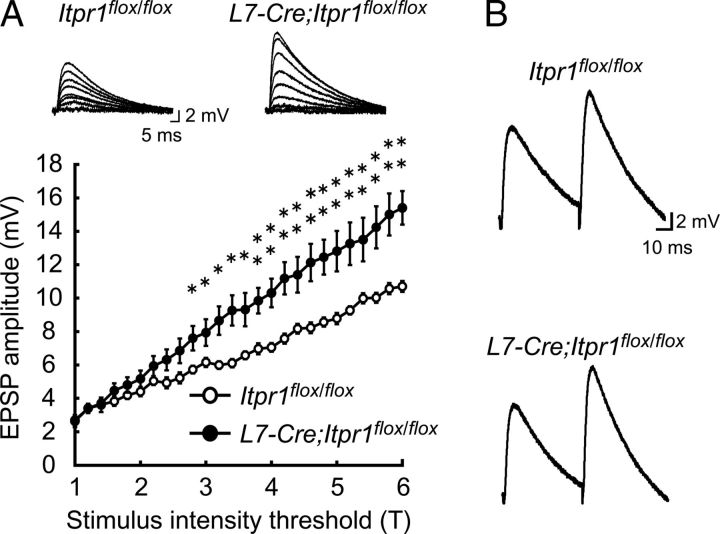
The structural maintenance of neural circuits is critical for higher brain functions in adulthood. Although several molecules have been identified as regulators for spine maintenance in hippocampal and cortical neurons, it is poorly understood how Purkinje cell (PC) spines are maintained in the mature cerebellum. Here we show that the calcium channel type 1 inositol trisphosphate receptor (IP3R1) in PCs plays a crucial role in controlling the maintenance of parallel fiber (PF)-PC synaptic circuits in the mature cerebellum in vivo. Significantly, adult mice lacking IP3R1 specifically in PCs (L7-Cre;Itpr1(flox/flox)) showed dramatic increase in spine density and spine length of PCs, despite having normal spines during development. In addition, the abnormally rearranged PF-PC synaptic circuits in mature cerebellum caused unexpectedly severe ataxia in adult L7-Cre;Itpr1(flox/flox) mice. Our findings reveal a specific role for IP3R1 in PCs not only as an intracellular mediator of cerebellar synaptic plasticity induction, but also as a critical regulator of PF-PC synaptic circuit maintenance in the mature cerebellum in vivo; this mechanism may underlie motor coordination and learning in adults.
Figures






References
-
- Arshavsky YI, Gelfand IM, Orlovsky GN. The cerebellum and control of rhythmical movements. Trends Neurosci. 1983;6:417–422. doi: 10.1016/0166-2236(83)90191-1. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases