

Slit/Robo signaling mediates spatial positioning of spiral ganglion neurons during development of cochlear innervation
- PMID: 23884932
- PMCID: PMC3721837
- DOI: 10.1523/JNEUROSCI.5736-12.2013
Slit/Robo signaling mediates spatial positioning of spiral ganglion neurons during development of cochlear innervation
Abstract
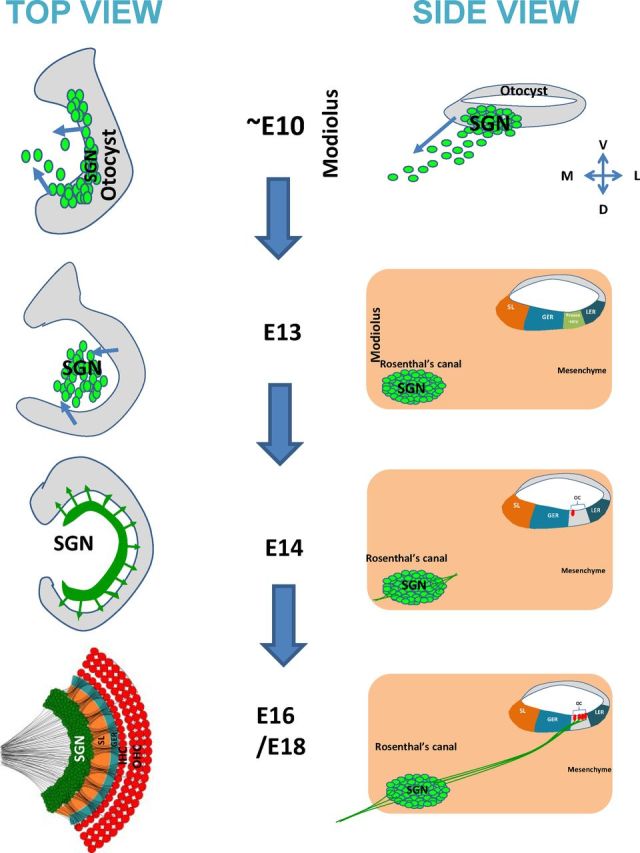
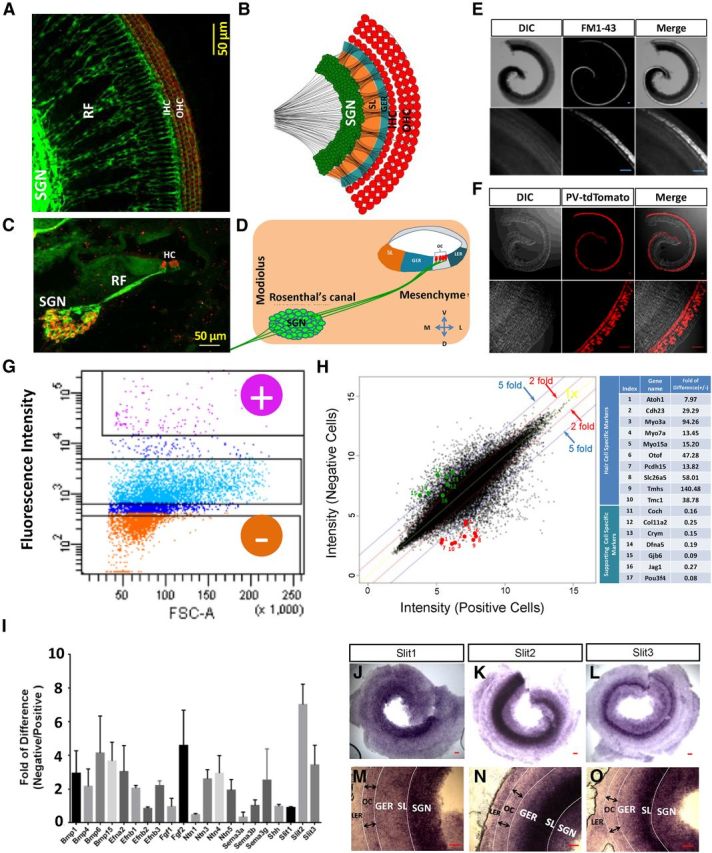
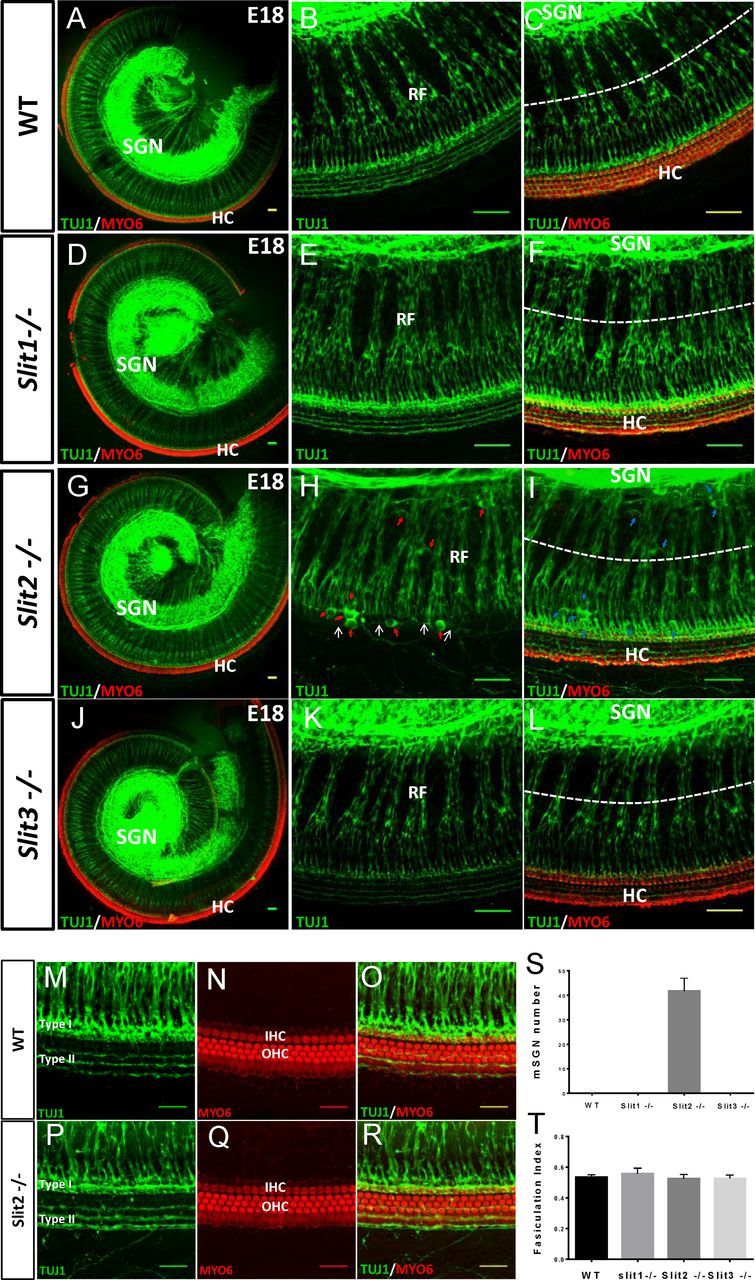
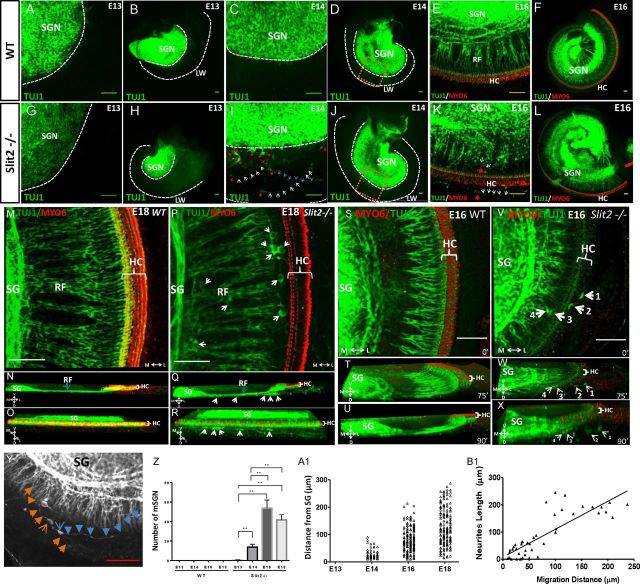
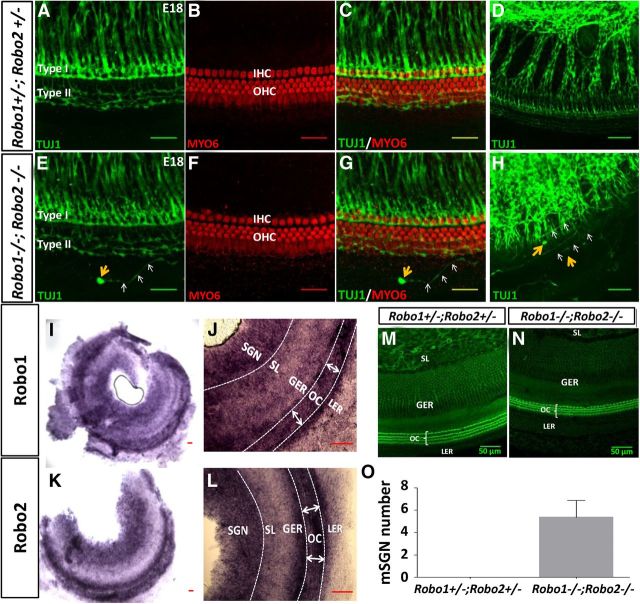
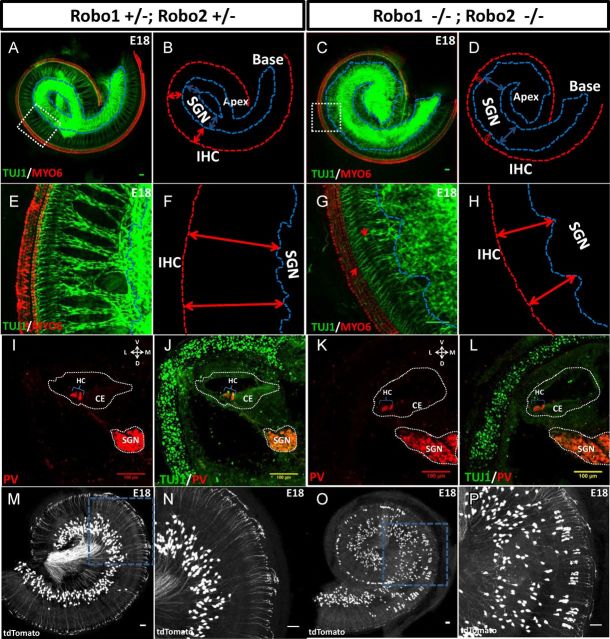
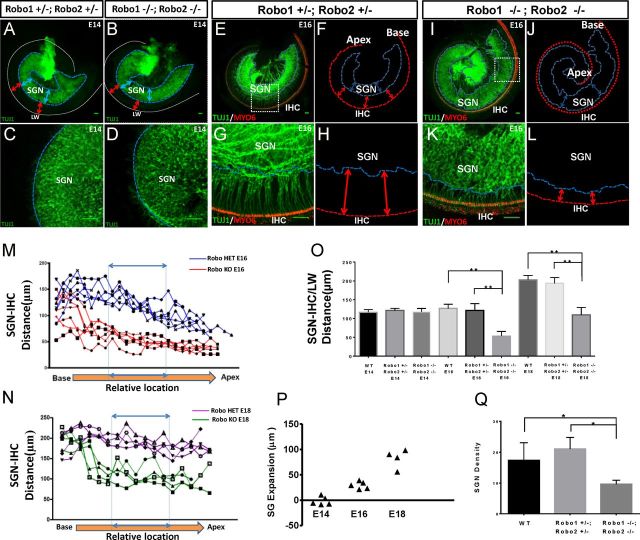
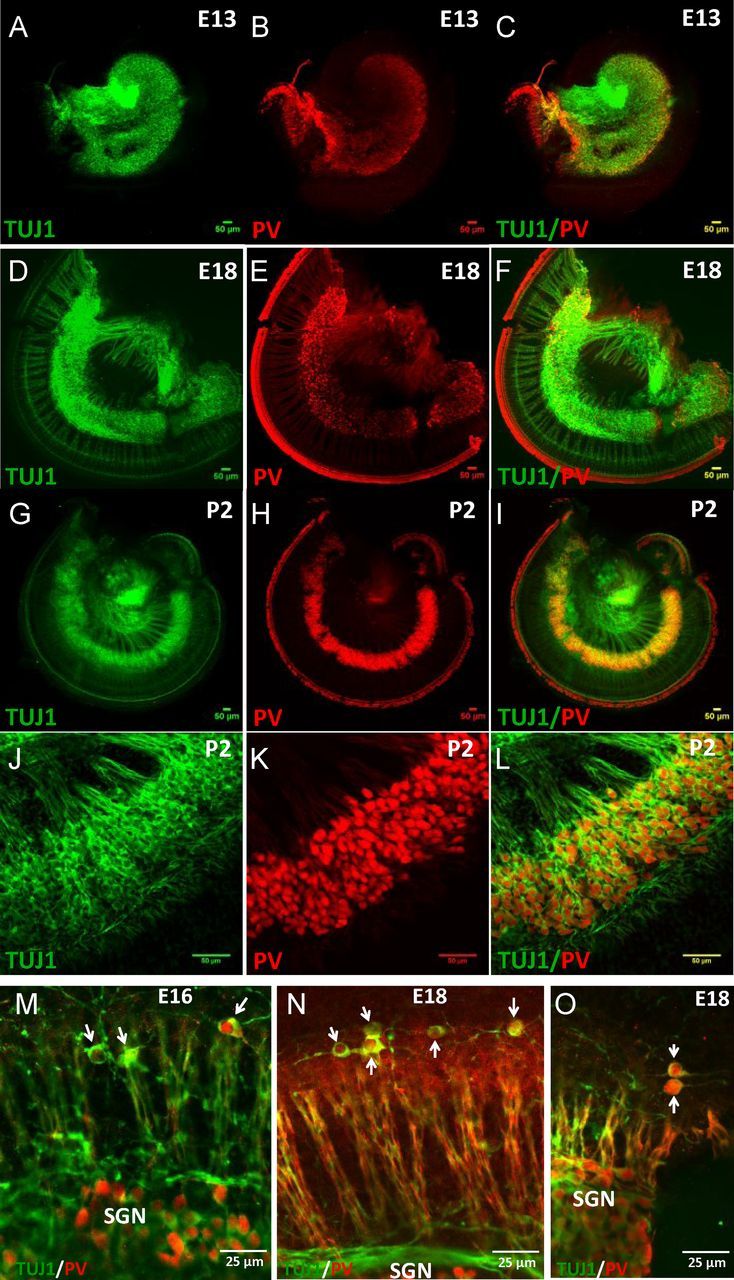
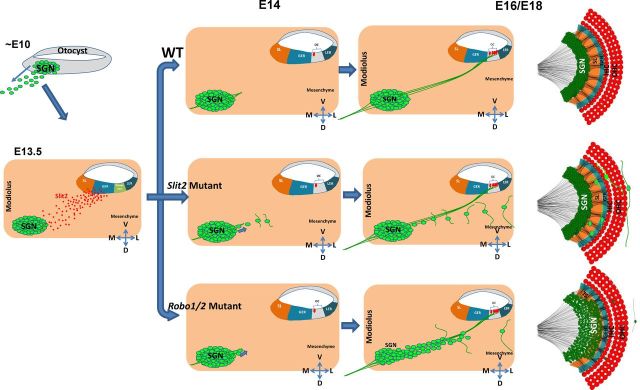
During the development of periphery auditory circuits, spiral ganglion neurons (SGNs) extend their neurites to innervate cochlear hair cells (HCs) with their soma aggregated into a cluster spatially segregated from the cochlear sensory epithelium. The molecular mechanisms underlying this spatial patterning remain unclear. In this study, in situ hybridization in the mouse cochlea suggests that Slit2 and its receptor, Robo1/2, exhibit apparently complementary expression patterns in the spiral ganglion and its nearby region, the spiral limbus. In Slit2 and Robo1/2 mutants, the spatial restriction of SGNs was disrupted. Mispositioned SGNs were found to scatter in the space between the cochlear epithelium and the main body of spiral ganglion, and the neurites of mispositioned SGNs were misrouted and failed to innervate HCs. Furthermore, in Robo1/2 mutants, SGNs were displaced toward the cochlear epithelium as an entirety. Examination of different embryonic stages in the mutants revealed that the mispositioning of SGNs was due to a progressive displacement to ectopic locations after their initial normal settlement at an earlier stage. Our results suggest that Slit/Robo signaling imposes a restriction force on SGNs to ensure their precise positioning for correct SGN-HC innervations.
Figures
References
-
- Borrell V, Cárdenas A, Ciceri G, Galcerán J, Flames N, Pla R, Nóbrega-Pereira S, García-Frigola C, Peregrín S, Zhao Z, Ma L, Tessier-Lavigne M, Marín O. Slit/Robo signaling modulates the proliferation of central nervous system progenitors. Neuron. 2012;76:338–352. doi: 10.1016/j.neuron.2012.08.003. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases