

Membrane peroxidation and methemoglobin formation are both necessary for band 3 clustering: mechanistic insights into human erythrocyte senescence
- PMID: 23889086
- PMCID: PMC3914982
- DOI: 10.1021/bi400405p
Membrane peroxidation and methemoglobin formation are both necessary for band 3 clustering: mechanistic insights into human erythrocyte senescence
Abstract
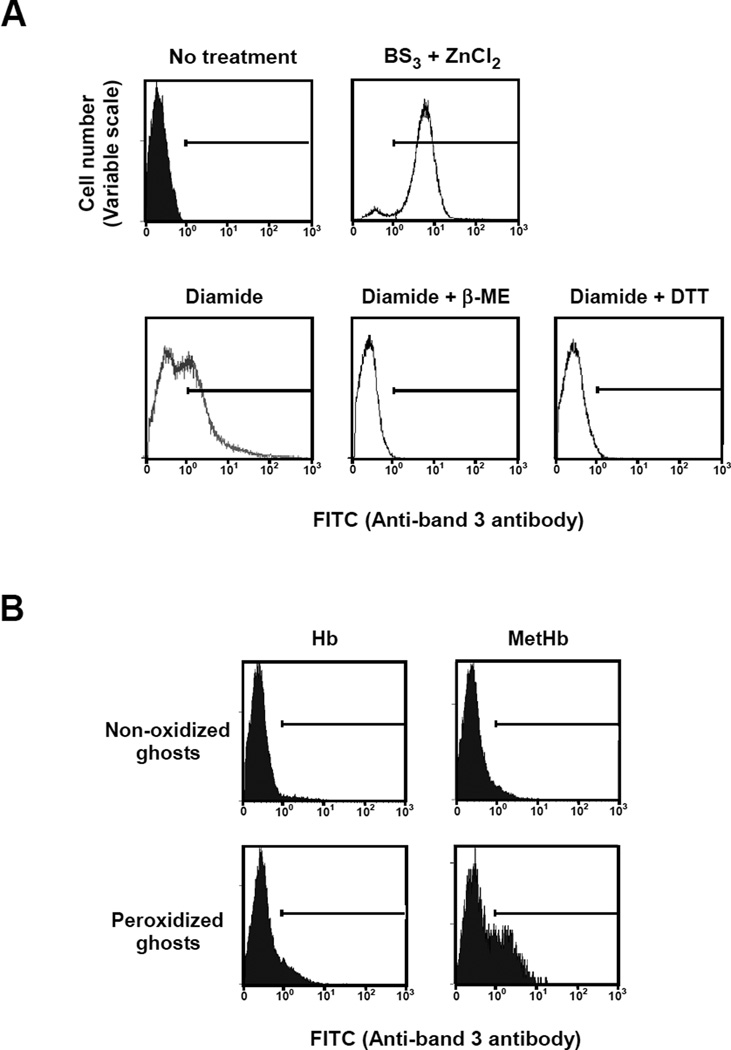
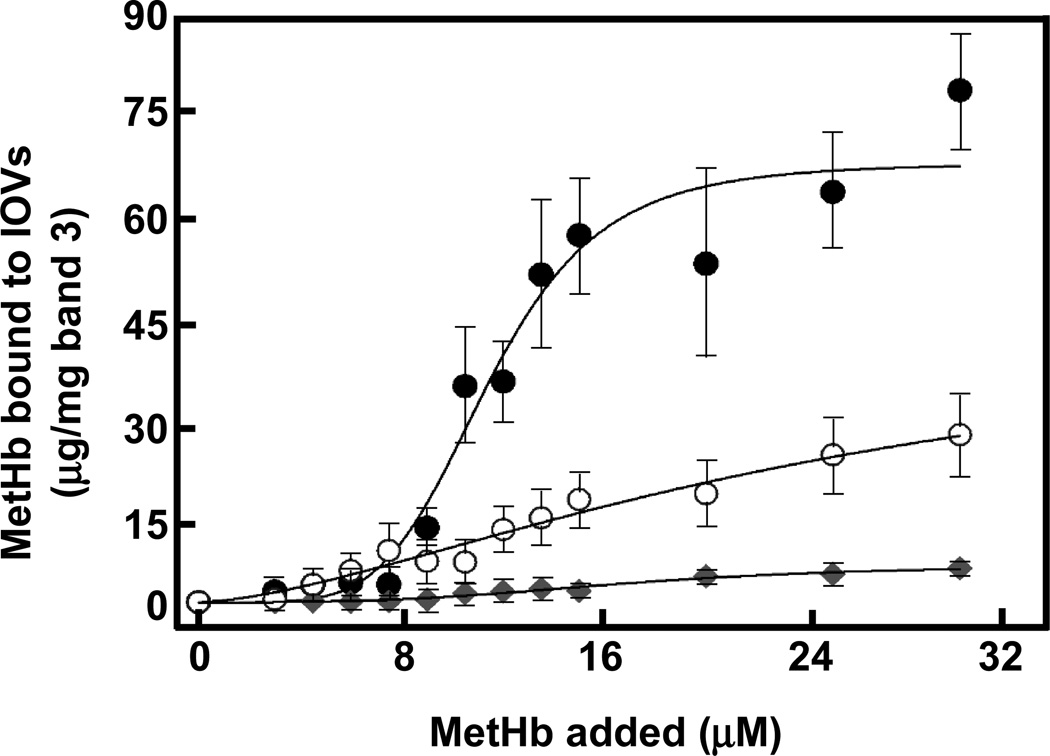
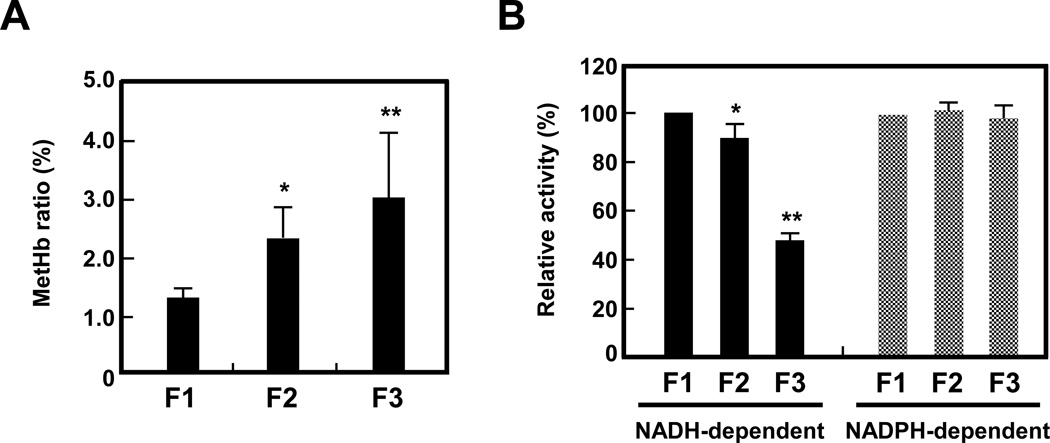
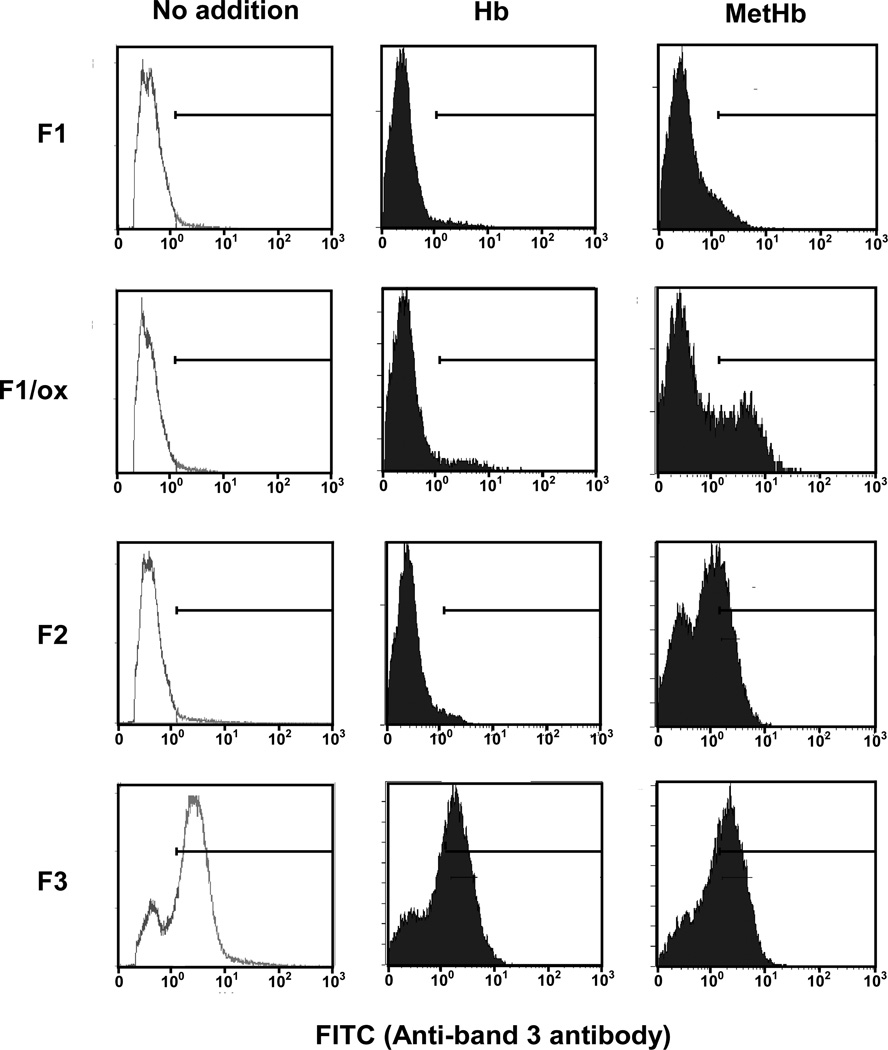
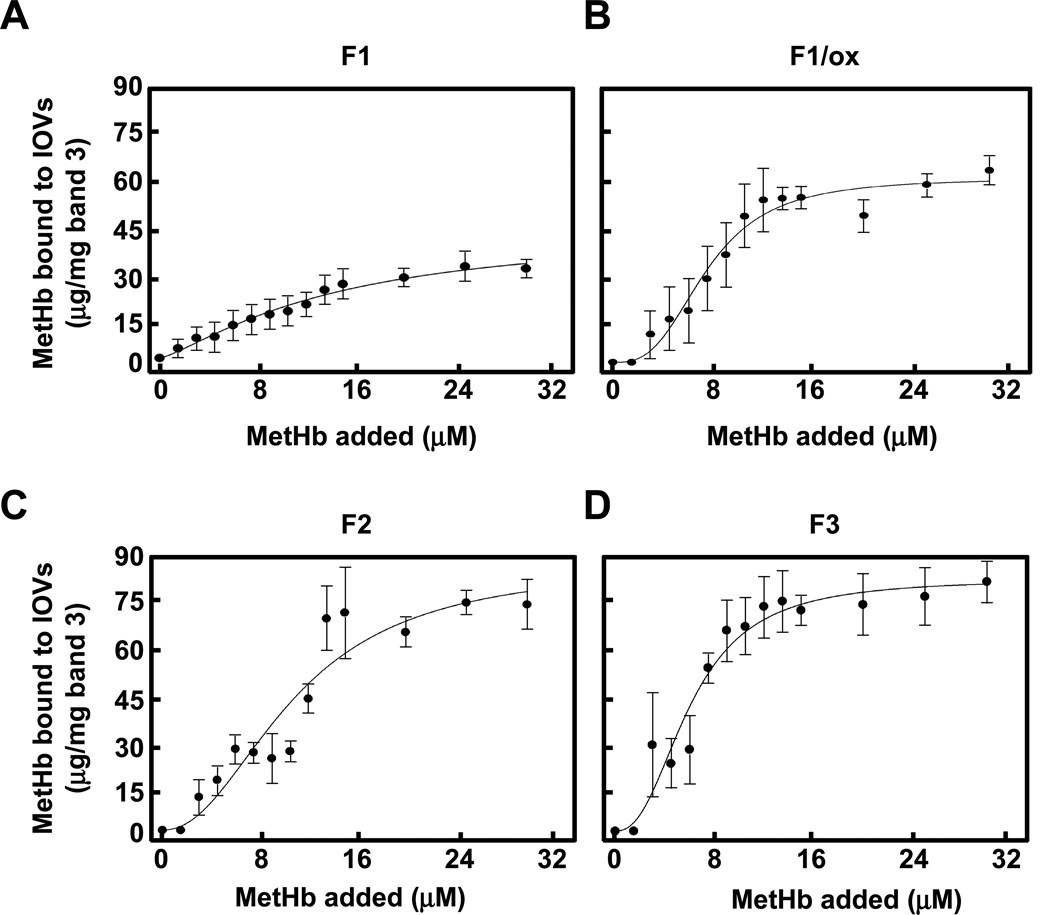
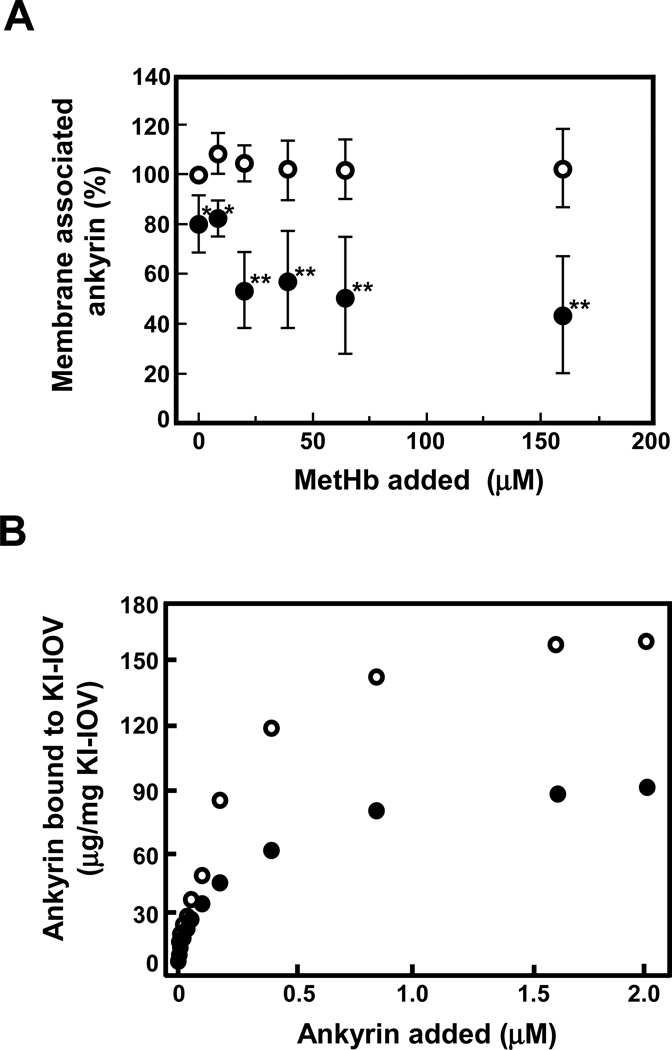
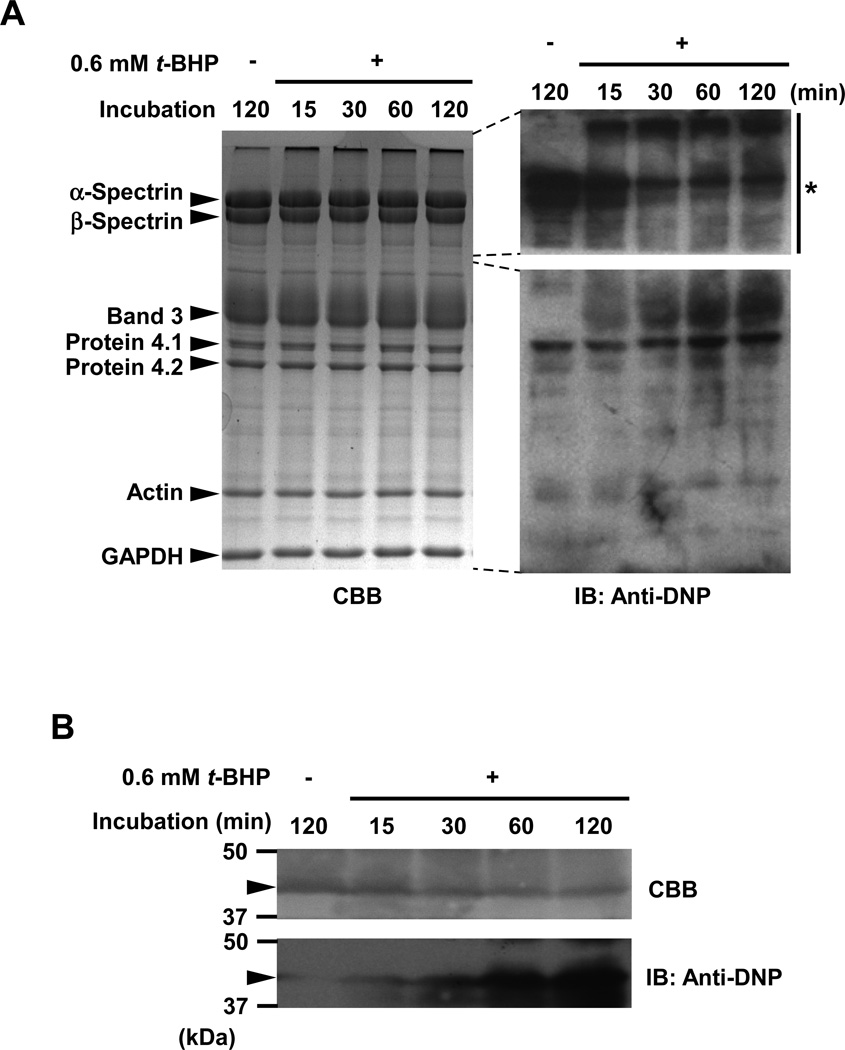
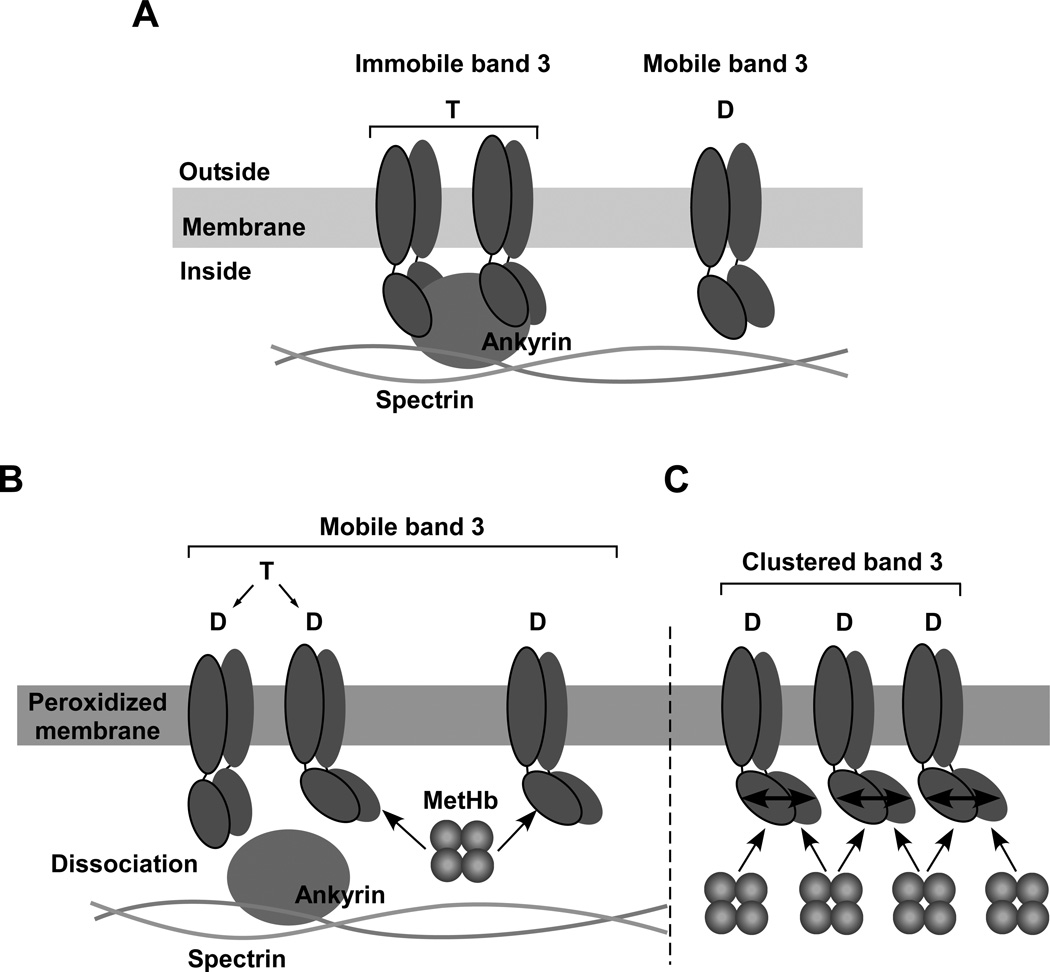
Oxidative damage and clustering of band 3 in the membrane have been implicated in the removal of senescent human erythrocytes from the circulation at the end of their 120 day life span. However, the biochemical and mechanistic events leading to band 3 cluster formation have yet to be fully defined. Here we show that while neither membrane peroxidation nor methemoglobin (MetHb) formation on their own can induce band 3 clustering in the human erythrocytes, they can do so when acting in combination. We further show that binding of MetHb to the cytoplasmic domain of band 3 in peroxidized, but not in untreated, erythrocyte membranes induces cluster formation. Age-fractionated populations of erythrocytes from normal human blood, obtained by a density gradient procedure, have allowed us to examine a subpopulation, highly enriched in senescent cells. We have found that band 3 clustering is a feature of only this small fraction, amounting to ∼0.1% of total circulating erythrocytes. These senescent cells are characterized by an increased proportion of MetHb as a result of reduced nicotinamide adenine dinucleotide-dependent reductase activity and accumulated oxidative membrane damage. These findings have allowed us to establish that the combined effects of membrane peroxidation and MetHb formation are necessary for band 3 clustering, and this is a very late event in erythrocyte life. A plausible mechanism for the combined effects of membrane peroxidation and MetHb is proposed, involving high-affinity cooperative binding of MetHb to the cytoplasmic domain of oxidized band 3, probably because of its carbonylation, rather than other forms of oxidative damage. This modification leads to dissociation of ankyrin from band 3, allowing the tetrameric MetHb to cross-link the resulting freely diffusible band 3 dimers, with formation of clusters.
Figures
References
-
- Burak Cimen MY. Free radical metabolism in human erythrocytes. Clinica. Chimica. Acta. 2008;390:1–11. - PubMed
-
- Signorini C, Ferrali M, Ciccoli L, Sugherini L, Magnani A, Comporti M. Iron release, membrane protein oxidation and erythrocyte ageing. FEBS Lett. 1995;362:165–170. - PubMed
-
- Hochstein P, Jain SK. Association of lipid peroxidation and polymerization of membrane proteins with erythrocyte aging. Fed. Proc. 1981;40:183–188. - PubMed
-
- Jain SK. Evidence for membrane lipid peroxidation during the in vivo aging of human erythrocytes. Biochim. Biophys. Acta. 1988;937:205–210. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
