

The expected value of control: an integrative theory of anterior cingulate cortex function
- PMID: 23889930
- PMCID: PMC3767969
- DOI: 10.1016/j.neuron.2013.07.007
The expected value of control: an integrative theory of anterior cingulate cortex function
Abstract
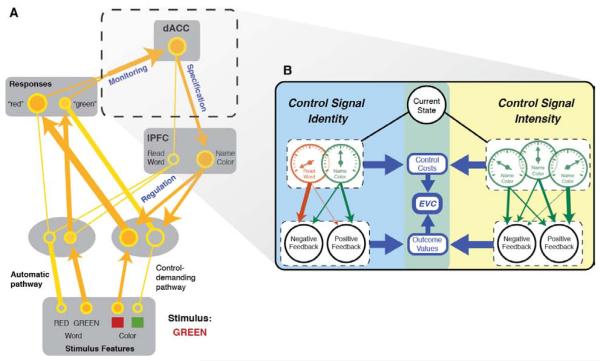
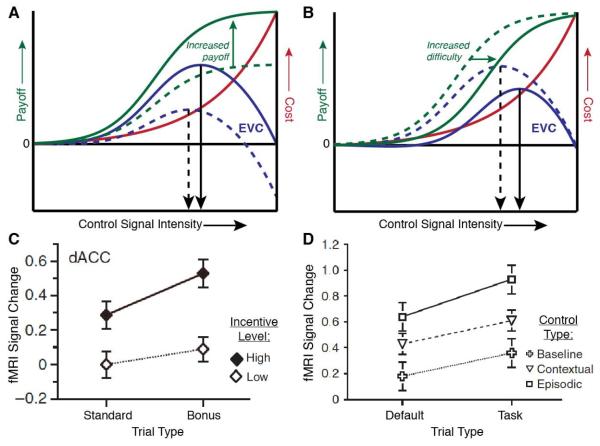
The dorsal anterior cingulate cortex (dACC) has a near-ubiquitous presence in the neuroscience of cognitive control. It has been implicated in a diversity of functions, from reward processing and performance monitoring to the execution of control and action selection. Here, we propose that this diversity can be understood in terms of a single underlying function: allocation of control based on an evaluation of the expected value of control (EVC). We present a normative model of EVC that integrates three critical factors: the expected payoff from a controlled process, the amount of control that must be invested to achieve that payoff, and the cost in terms of cognitive effort. We propose that dACC integrates this information, using it to determine whether, where and how much control to allocate. We then consider how the EVC model can explain the diverse array of findings concerning dACC function.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Alexander MP, Stuss DT, Picton T, Shallice T, Gillingham S. Regional frontal injuries cause distinct impairments in cognitive control. Neurology. 2007;68:1515–1523. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
