

A network for scene processing in the macaque temporal lobe
- PMID: 23891401
- PMCID: PMC8127731
- DOI: 10.1016/j.neuron.2013.06.015
A network for scene processing in the macaque temporal lobe
Abstract
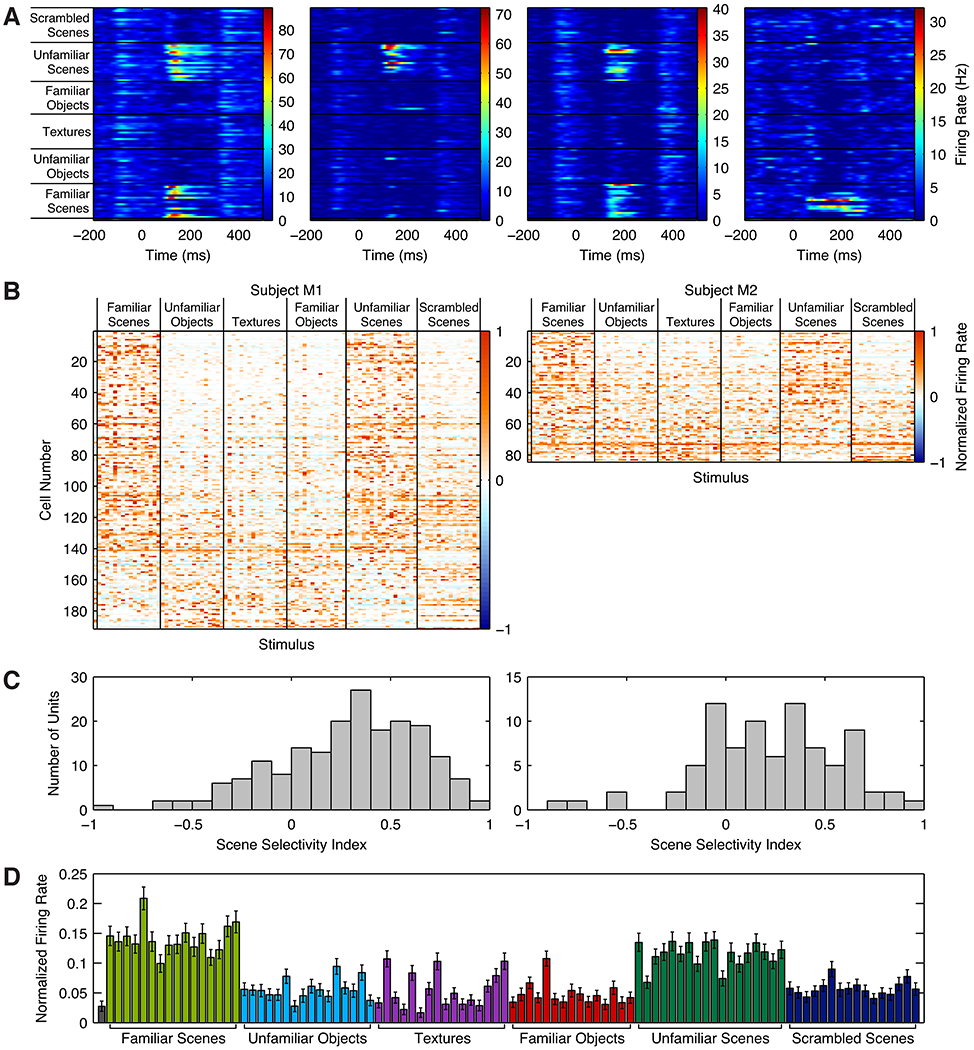
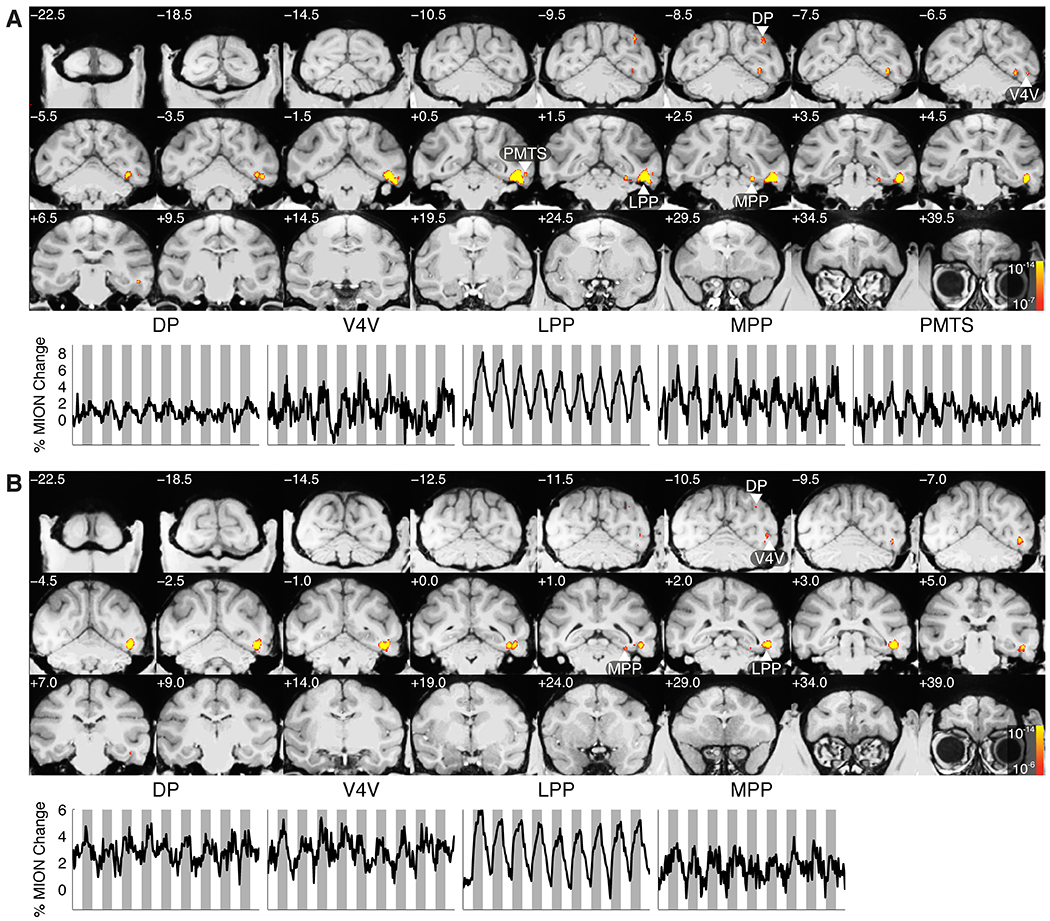
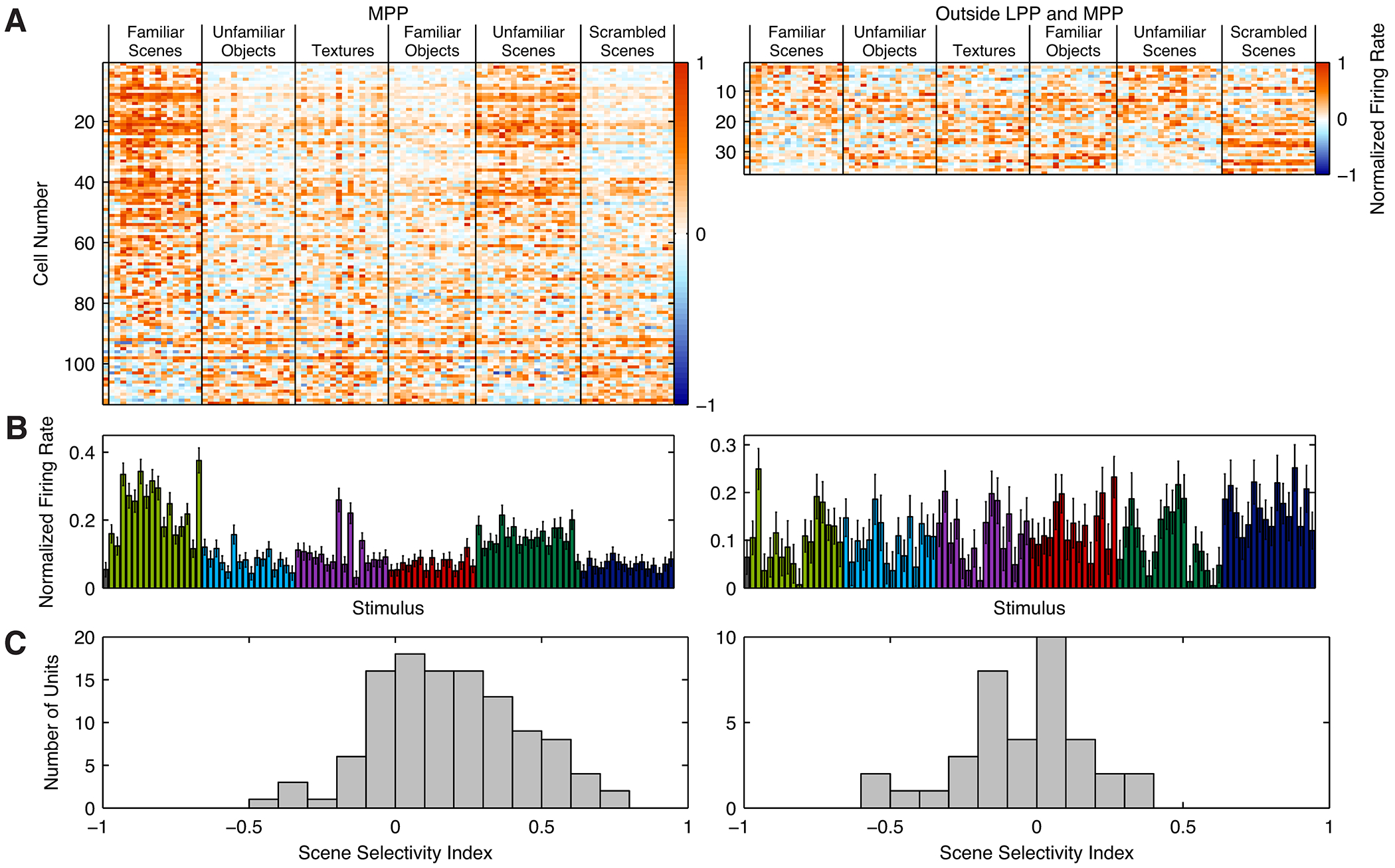
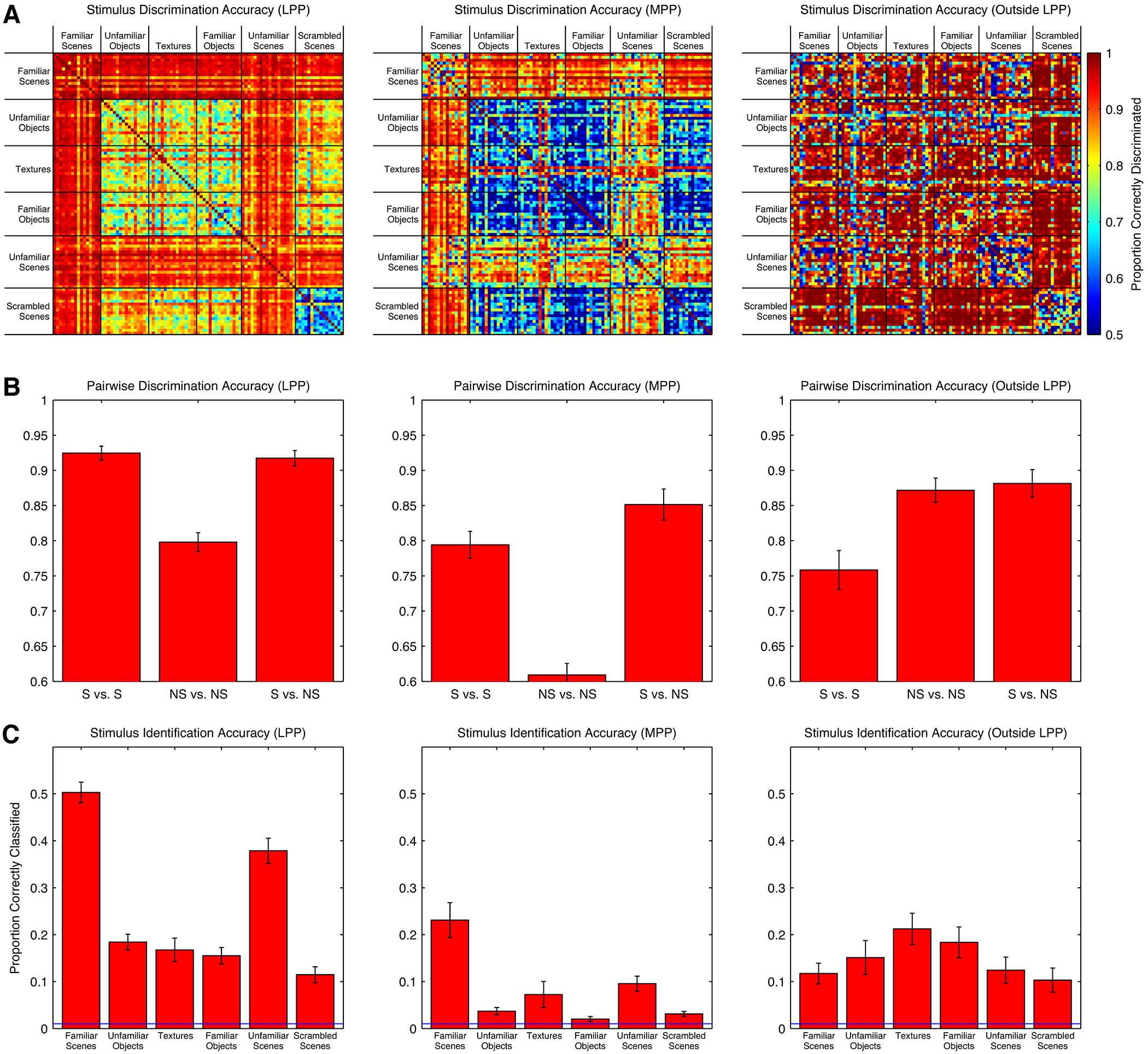
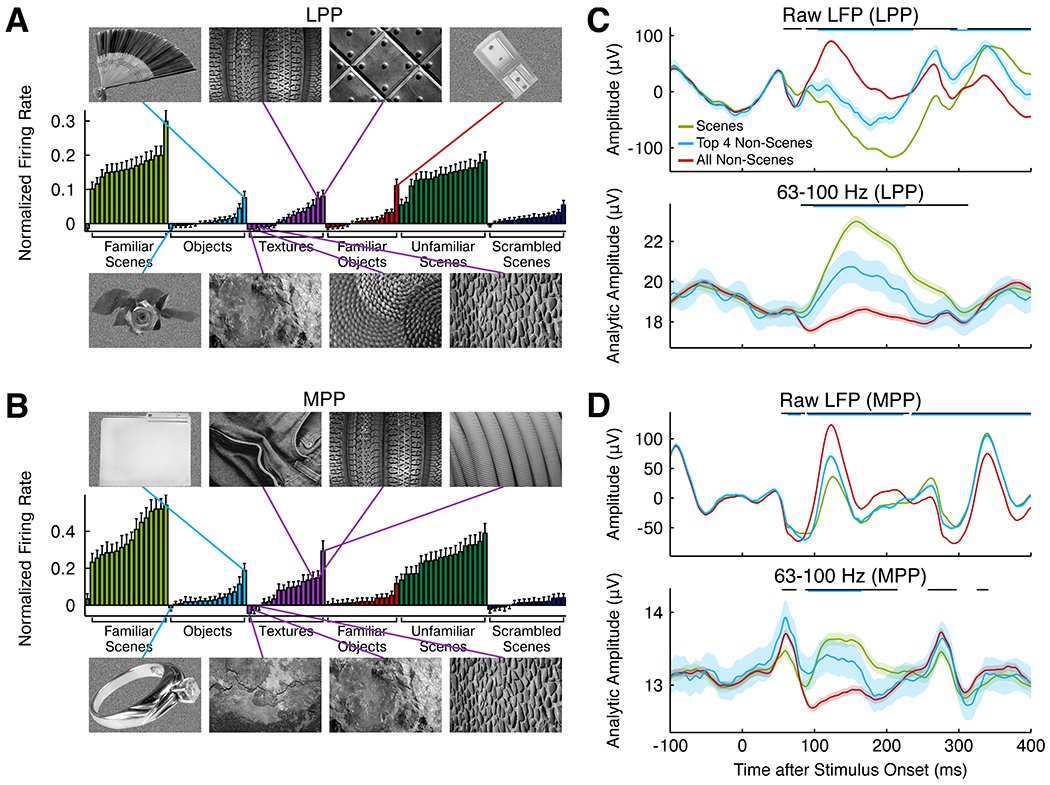
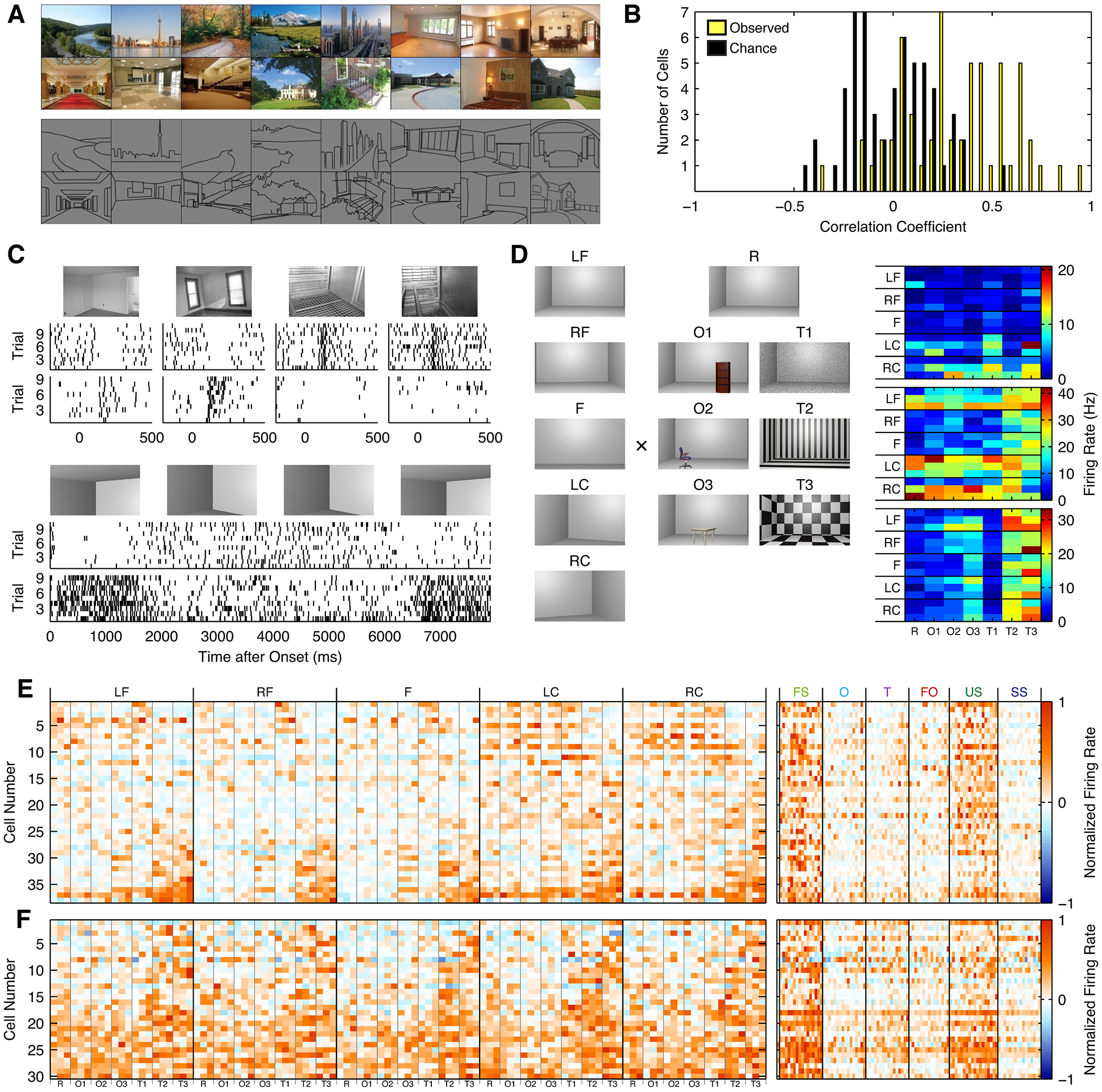
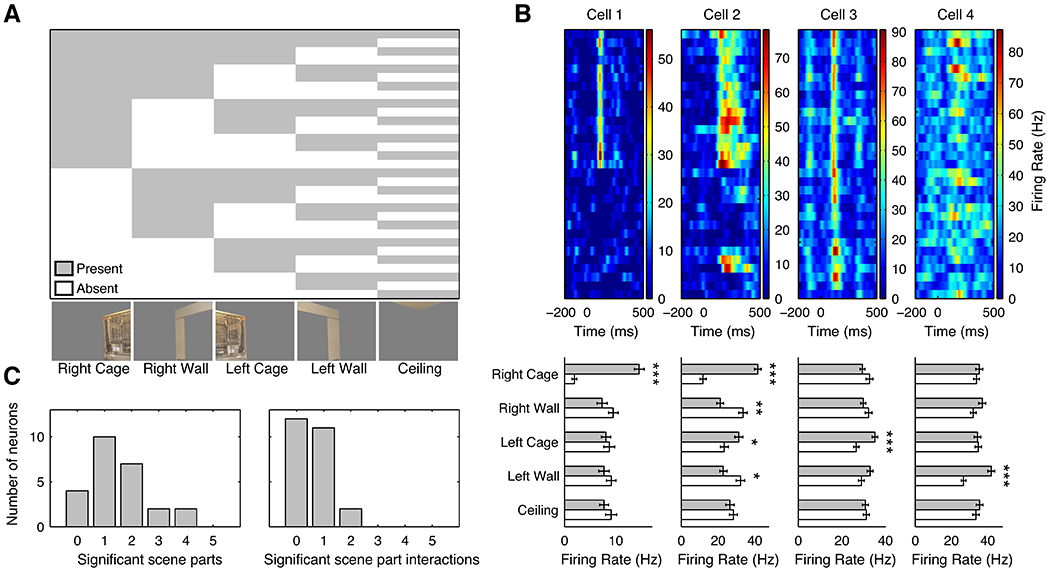
Spatial navigation is a complex process, but one that is essential for any mobile organism. We localized a region in the macaque occipitotemporal sulcus that responds preferentially to images of scenes. Single-unit recording revealed that this region, which we term the lateral place patch (LPP), contained a large concentration of scene-selective single units. These units were not modulated by spatial layout alone but were instead modulated by a combination of spatial and nonspatial factors, with individual units coding specific scene parts. We further demonstrate by microstimulation that LPP is connected with extrastriate visual areas V4V and DP and a scene-selective medial place patch in the parahippocampal gyrus, revealing a ventral network for visual scene processing in the macaque.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Scene areas in humans and macaques.Neuron. 2013 Aug 21;79(4):615-7. doi: 10.1016/j.neuron.2013.08.001. Neuron. 2013. PMID: 23972591 Free PMC article.
References
-
- Aguirre GK, and D’Esposito M (1999). Topographical disorientation: a synthesis and taxonomy. Brain 122, 1613. - PubMed
-
- Blatt GJ, and Rosene DL (1998). Organization of direct hippocampal efferent projections to the cerebral cortex of the rhesus monkey: Projections from CA1, prosubiculum, and subiculum to the temporal lobe. J. Comp. Neurol 392, 92–114. - PubMed
-
- Blatt GJ, Pandya DN, and Rosene DL (2003). Parcellation of cortical afferents to three distinct sectors in the parahippocampal gyrus of the rhesus monkey: An anatomical and neurophysiological study. J. Comp. Neurol 466, 161–179. - PubMed
-
- Boussaoud D, Desimone R, and Ungerleider LG (1991). Visual topography of area TEO in the macaque. J. Comp. Neurol 306, 554–575. - PubMed
-
- Buhrmester M, Kwang T, and Gosling SD (2011). Amazon’s Mechanical Turk: A new source of inexpensive, yet high-quality, data? Perspect. Psychol. Sci 6, 3–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
