

Functional loss of two ceramide synthases elicits autophagy-dependent lifespan extension in C. elegans
- PMID: 23894595
- PMCID: PMC3716707
- DOI: 10.1371/journal.pone.0070087
Functional loss of two ceramide synthases elicits autophagy-dependent lifespan extension in C. elegans
Abstract
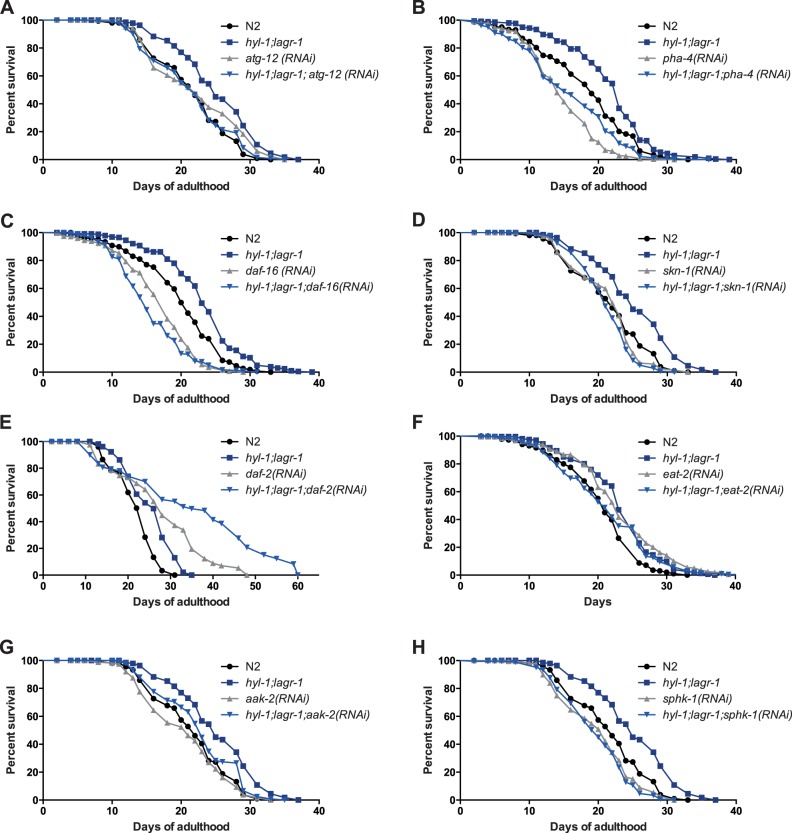
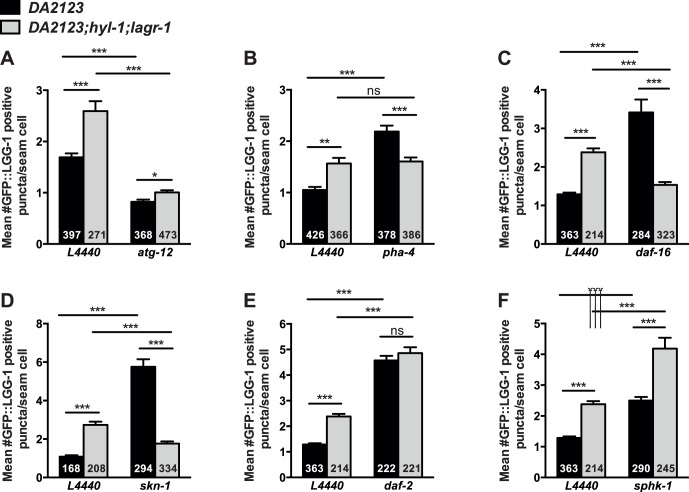
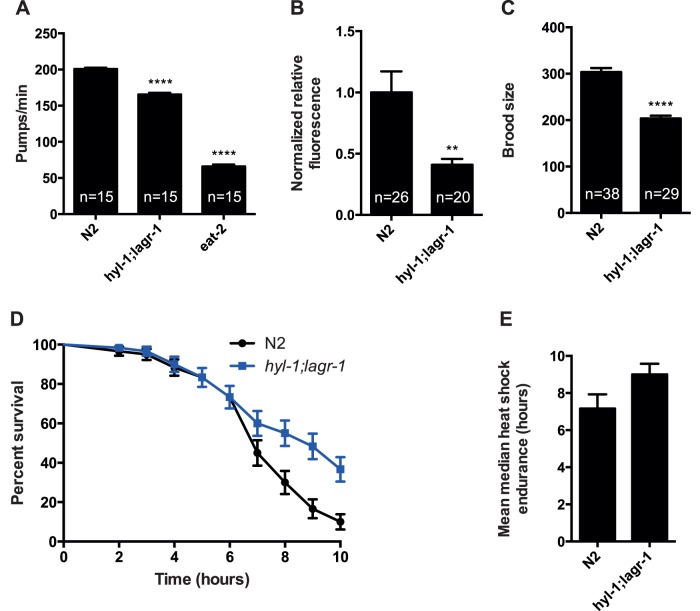
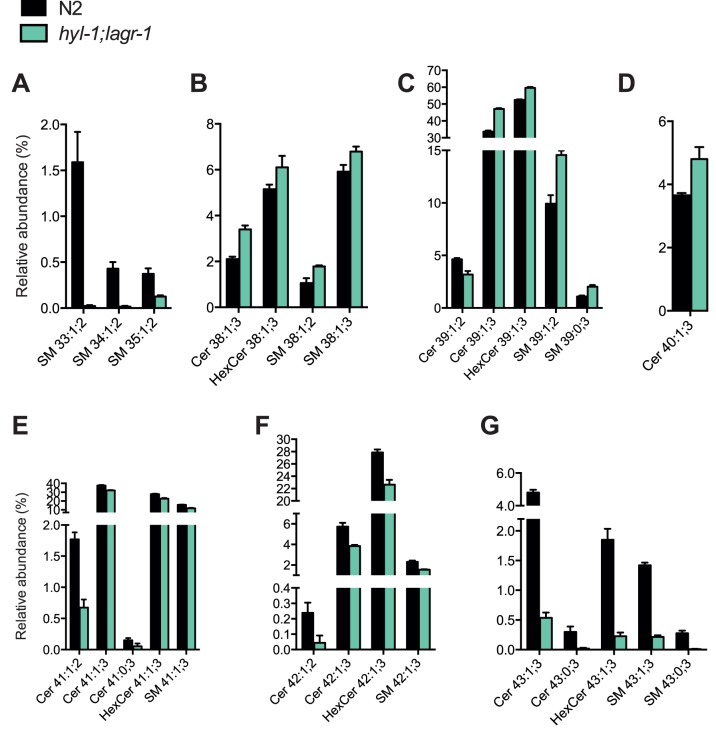
Ceramide and its metabolites constitute a diverse group of lipids, which play important roles as structural entities of biological membranes as well as regulators of cellular growth, differentiation, and development. The C. elegans genome comprises three ceramide synthase genes; hyl-1, hyl-2, and lagr-1. HYL-1 function is required for synthesis of ceramides and sphingolipids containing very long acyl-chains (≥C24), while HYL-2 is required for synthesis of ceramides and sphingolipids containing shorter acyl-chains (≤C22). Here we show that functional loss of HYL-2 decreases lifespan, while loss of HYL-1 or LAGR-1 does not affect lifespan. We show that loss of HYL-1 and LAGR-1 functions extend lifespan in an autophagy-dependent manner, as knock down of the autophagy-associated gene ATG-12 abolishes hyl-1;lagr-1 longevity. The transcription factors PHA-4/FOXA, DAF-16/FOXO, and SKN-1 are also required for the observed lifespan extension, as well as the increased number of autophagosomes in hyl-1;lagr-1 animals. Both autophagic events and the transcription factors PHA-4/FOXA, DAF-16, and SKN-1 have previously been associated with dietary restriction-induced longevity. Accordingly, we find that hyl-1;lagr-1 animals display reduced feeding, increased resistance to heat, and reduced reproduction. Collectively, our data suggest that specific sphingolipids produced by different ceramide synthases have opposing roles in determination of C. elegans lifespan. We propose that loss of HYL-1 and LAGR-1 result in dietary restriction-induced autophagy and consequently prolonged longevity.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
