

Sleep fragmentation promotes NADPH oxidase 2-mediated adipose tissue inflammation leading to insulin resistance in mice
- PMID: 23897221
- PMCID: PMC3907464
- DOI: 10.1038/ijo.2013.139
Sleep fragmentation promotes NADPH oxidase 2-mediated adipose tissue inflammation leading to insulin resistance in mice
Abstract
Background: Short sleep has been implicated in higher risk of obesity in humans, and is associated with insulin resistance. However, the effects of fragmented sleep (SF) rather than curtailed sleep on glucose homeostasis are unknown.
Methods: Wild-type and NADPH oxidase 2 (Nox2) null male mice were subjected to SF or sleep control conditions for 3 days to 3 weeks. Systemic and visceral adipose tissue (VAT) insulin sensitivity tests, glucose tolerance test, fluorescence-activated cell sorting and immunohistochemistry for macrophages and its sub-types (M1 and M2), and Nox expression and activity were examined.
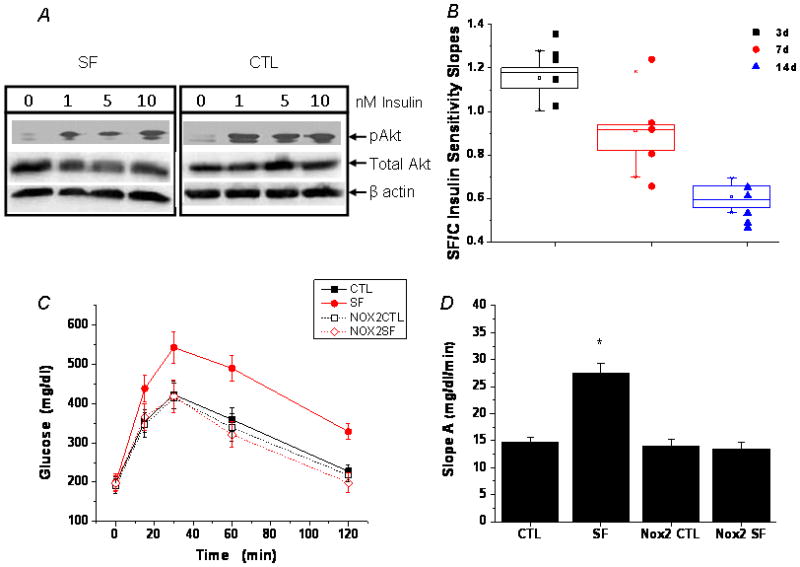
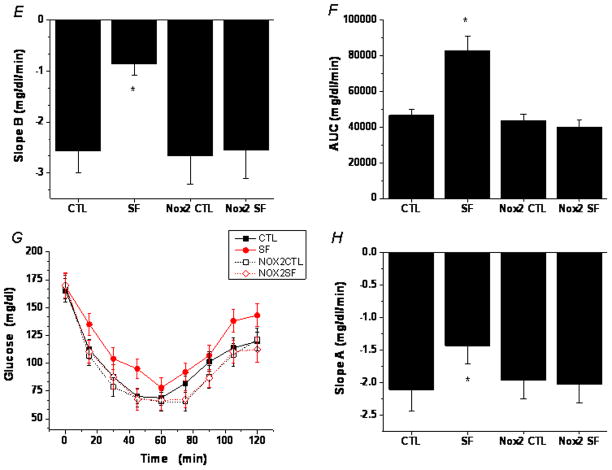
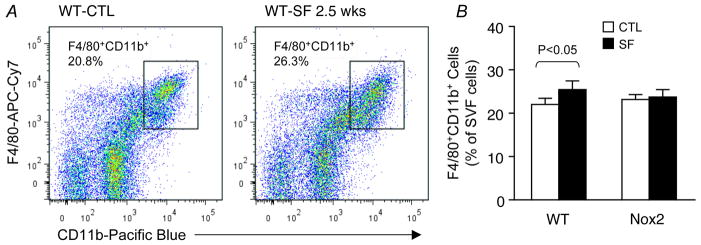
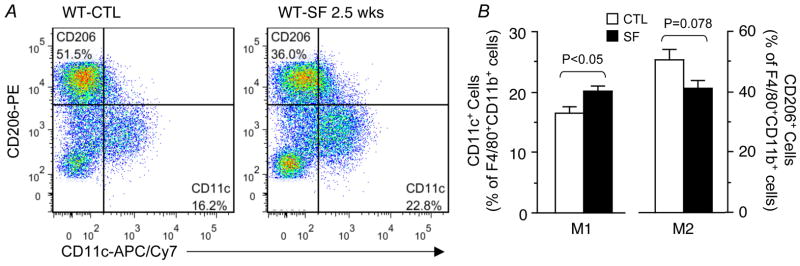
Results: Here we show that SF in the absence of sleep curtailment induces time-dependent insulin resistance, in vivo and also in vitro in VAT. Oxidative stress pathways were upregulated by SF in VAT, and were accompanied by M1 macrophage polarization. SF-induced oxidative stress, inflammation and insulin resistance in VAT were completely abrogated in genetically altered mice lacking Nox2 activity.
Conclusions: These studies imply that SF, a frequent occurrence in many disorders and more specifically in sleep apnea, is a potent inducer of insulin resistance via activation of oxidative stress and inflammatory pathways, thereby opening the way for therapeutic strategies.
Conflict of interest statement
Figures
References
-
- Spiegel K, Leproult R, Van Cauter E. Impact of sleep debt on metabolic and endocrine function. Lancet. 1999;354:1435–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
