

Interleukin-25 (IL-25) promotes efficient protective immunity against Trichinella spiralis infection by enhancing the antigen-specific IL-9 response
- PMID: 23897610
- PMCID: PMC3811766
- DOI: 10.1128/IAI.00646-13
Interleukin-25 (IL-25) promotes efficient protective immunity against Trichinella spiralis infection by enhancing the antigen-specific IL-9 response
Abstract
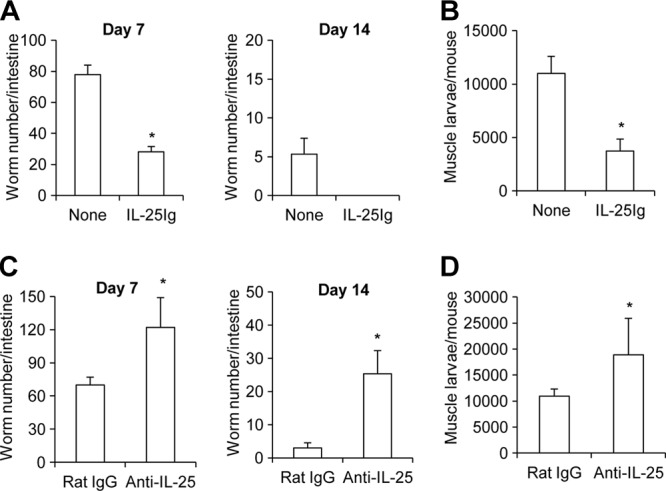
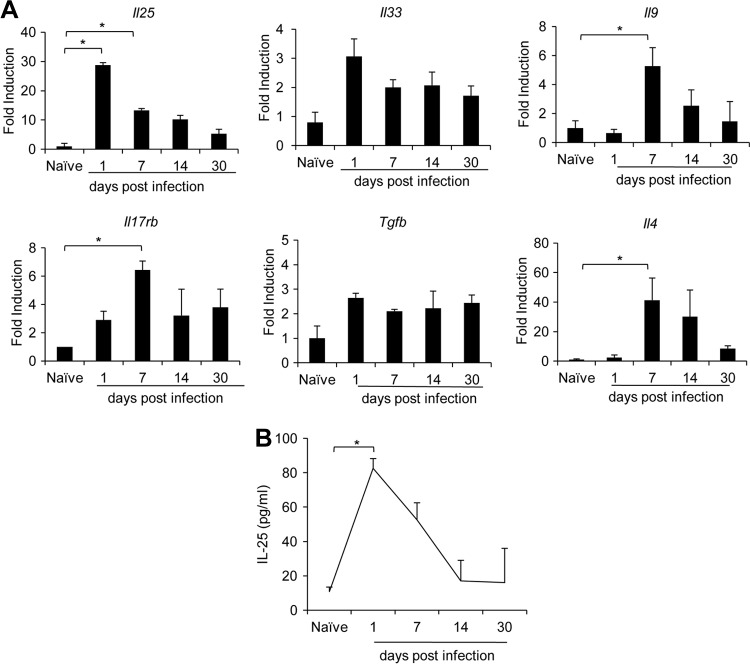
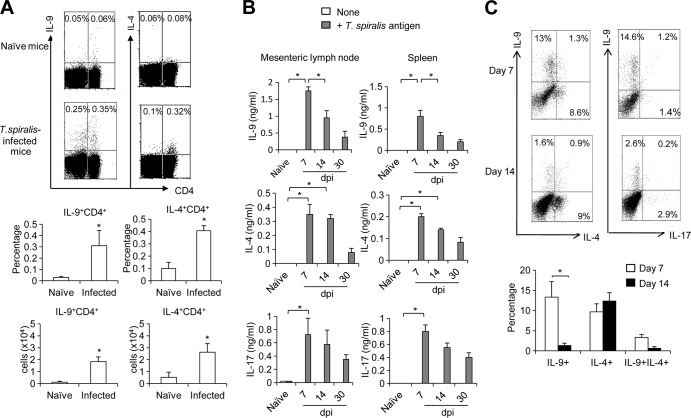
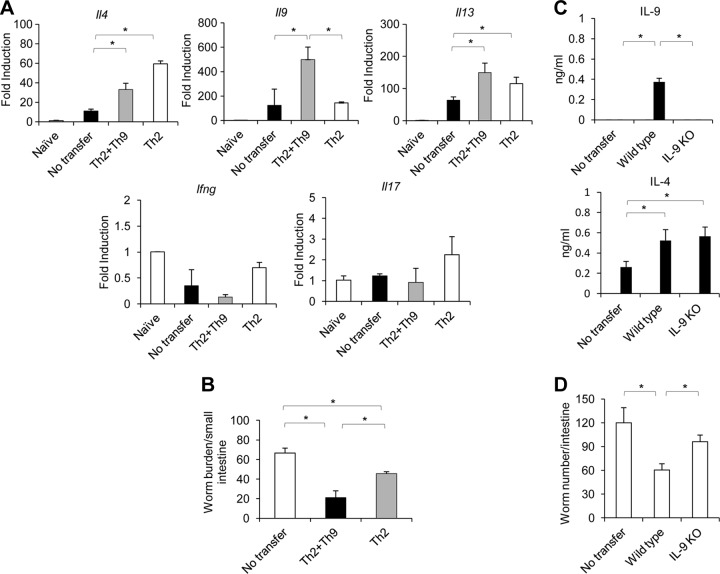
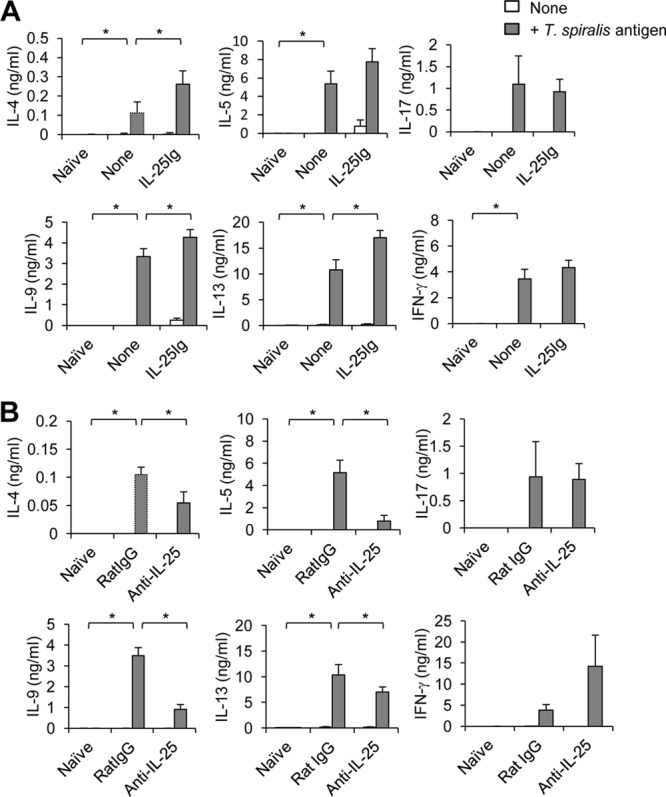
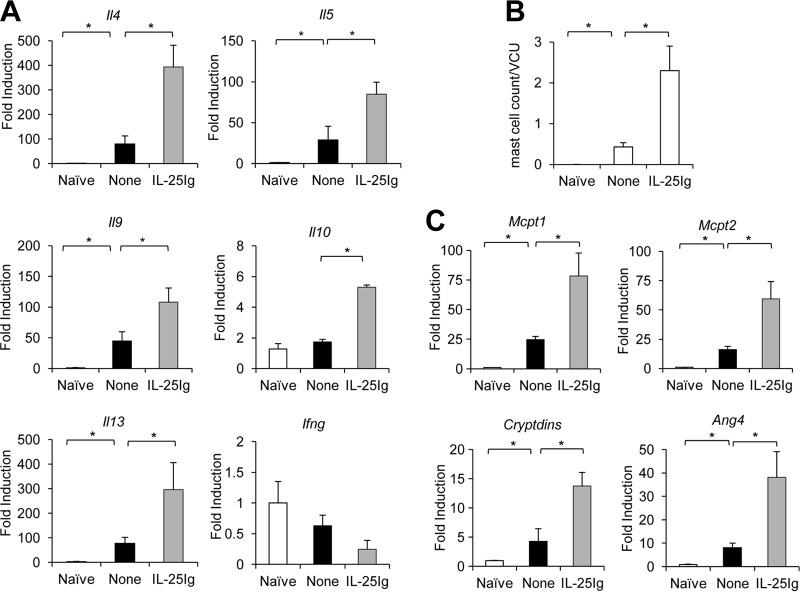
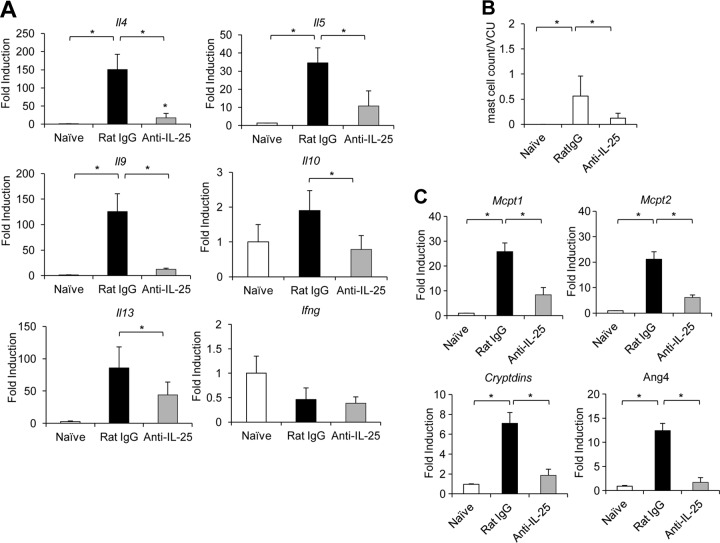
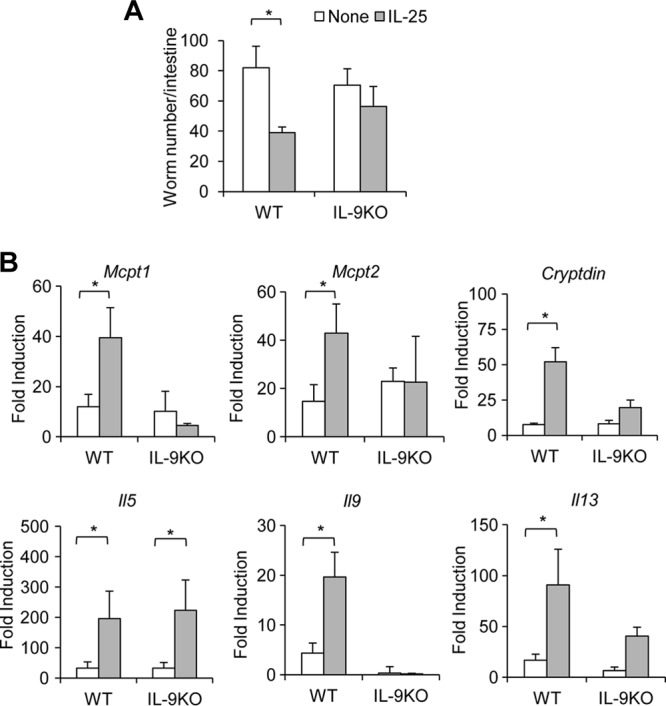
Mammalian hosts often develop distinct immune response against the diverse parasitic helminths that have evolved for immune evasion. Interleukin-25 (IL-25), an IL-17 cytokine family member, plays a key role in initiating the protective immunity against several parasitic helminths; however, the involvement and underlying mechanisms by which IL-25 mediates immune response against Trichinella spiralis infection have not been investigated. Here we showed that IL-25 functions in promoting protective immunity against T. spiralis infection. Mice treated with IL-25 exhibited a lower worm burden and fewer muscle larvae in the later stage of T. spiralis infection. In contrast, mice treated with neutralizing antibody against IL-25 failed to expel T. spiralis effectively. During T. spiralis infection, intestinal IL-25 expression was rapidly elevated before the onset of IL-4 and IL-9 induction. While antigen-specific Th2 and Th9 immune responses were both developed during T. spiralis infection, an antigen-specific Th9 response appeared to be transiently induced in the early stage of infection. Mice into which antigen-specific T cells deficient in IL-9 were transferred were less effective in worm clearance than those given wild-type T cells. The strength of the antigen-specific Th9 immune response against T. spiralis could be enhanced or attenuated after treatment with IL-25 or neutralizing antibody against IL-25, respectively, correlating positively with the levels of intestinal mastocytosis and the expression of IL-9-regulated genes, including mast cell- and Paneth cell-specific genes. Thus, our study demonstrates that intestinal IL-25 promotes protective immunity against T. spiralis infection by inducing antigen-specific Th9 immune response.
Figures
References
-
- Finkelman FD, Shea-Donohue T, Goldhill J, Sullivan CA, Morris SC, Madden KB, Gause WC, Urban JF., Jr 1997. Cytokine regulation of host defense against parasitic gastrointestinal nematodes: lessons from studies with rodent models. Annu. Rev. Immunol. 15:505–533 - PubMed
-
- Else KJ, Finkelman FD. 1998. Intestinal nematode parasites, cytokines and effector mechanisms. Int. J. Parasitol. 28:1145–1158 - PubMed
-
- Fort MM, Cheung J, Yen D, Li J, Zurawski SM, Lo S, Menon S, Clifford T, Hunte B, Lesley R, Muchamuel T, Hurst SD, Zurawski G, Leach MW, Gorman DM, Rennick DM. 2001. IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 15:985–995 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
