

Reprogramming cells and tissue patterning via bioelectrical pathways: molecular mechanisms and biomedical opportunities
- PMID: 23897652
- PMCID: PMC3841289
- DOI: 10.1002/wsbm.1236
Reprogramming cells and tissue patterning via bioelectrical pathways: molecular mechanisms and biomedical opportunities
Abstract
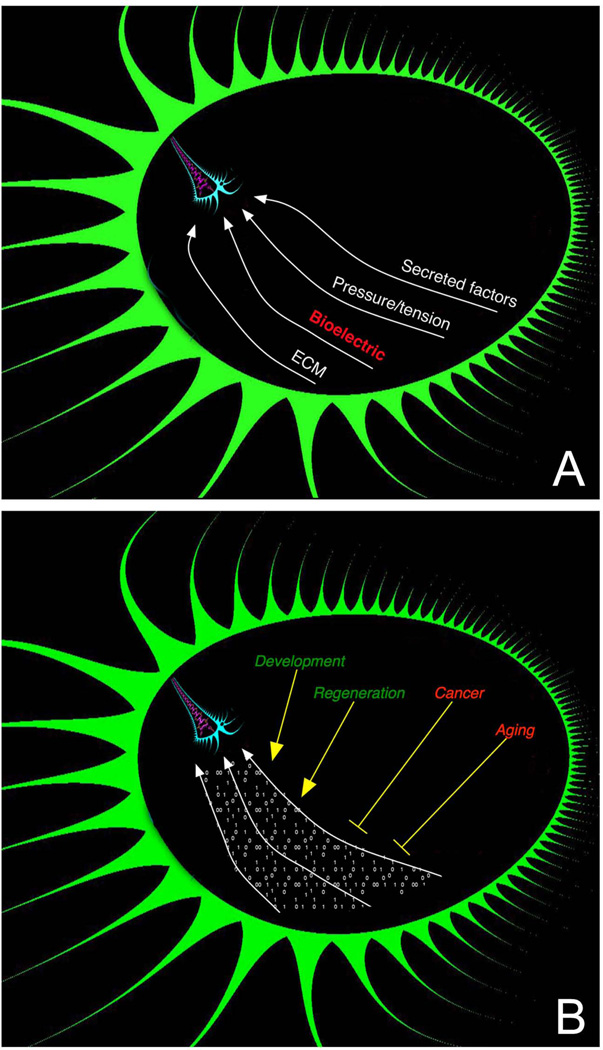
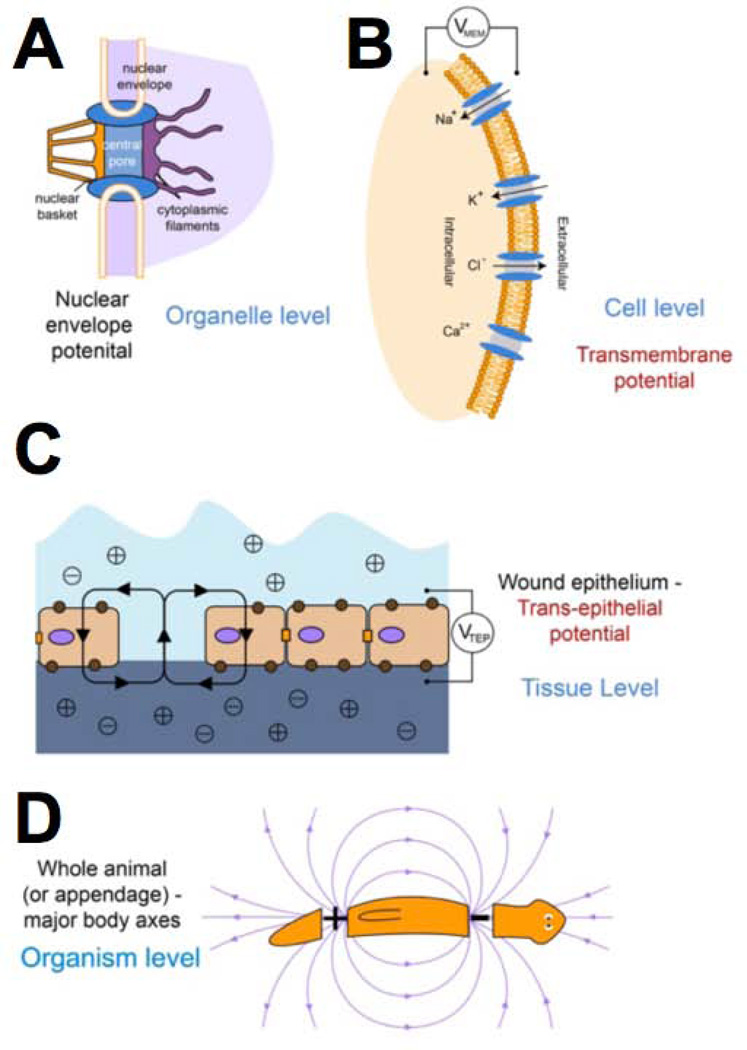
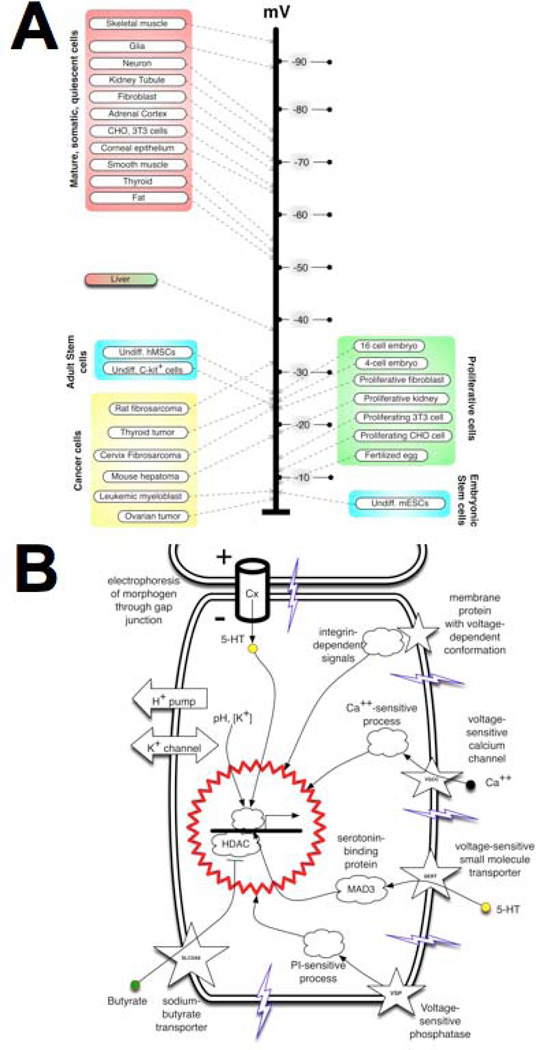
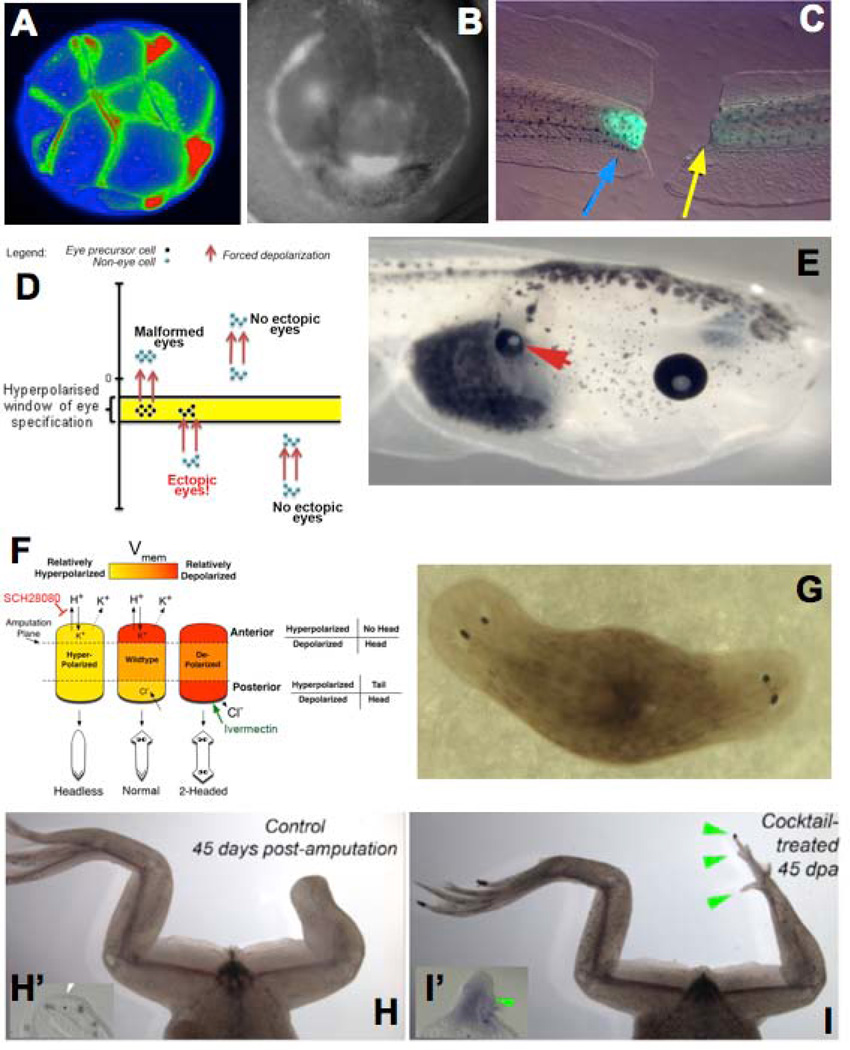
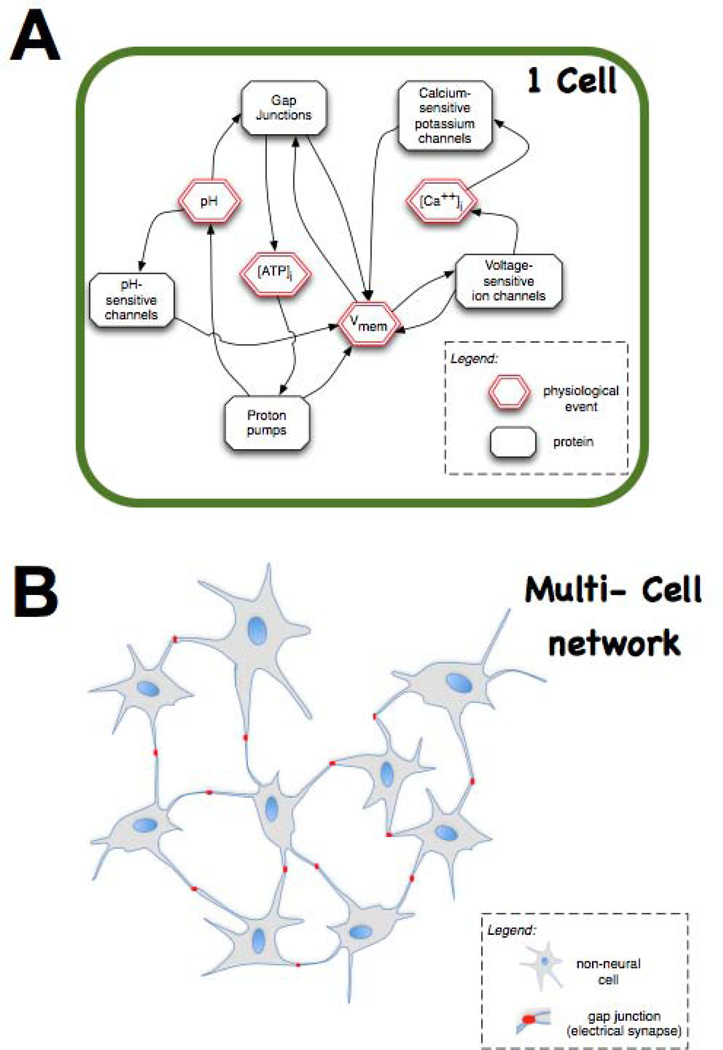
Transformative impact in regenerative medicine requires more than the reprogramming of individual cells: advances in repair strategies for birth defects or injuries, tumor normalization, and the construction of bioengineered organs and tissues all require the ability to control large-scale anatomical shape. Much recent work has focused on the transcriptional and biochemical regulation of cell behavior and morphogenesis. However, exciting new data reveal that bioelectrical properties of cells and their microenvironment exert a profound influence on cell differentiation, proliferation, and migration. Ion channels and pumps expressed in all cells, not just excitable nerve and muscle, establish resting potentials that vary across tissues and change with significant developmental events. Most importantly, the spatiotemporal gradients of these endogenous transmembrane voltage potentials (Vmem ) serve as instructive patterning cues for large-scale anatomy, providing organ identity, positional information, and prepattern template cues for morphogenesis. New genetic and pharmacological techniques for molecular modulation of bioelectric gradients in vivo have revealed the ability to initiate complex organogenesis, change tissue identity, and trigger regeneration of whole vertebrate appendages. A large segment of the spatial information processing that orchestrates individual cells' programs toward the anatomical needs of the host organism is electrical; this blurs the line between memory and decision-making in neural networks and morphogenesis in nonneural tissues. Advances in cracking this bioelectric code will enable the rational reprogramming of shape in whole tissues and organs, revolutionizing regenerative medicine, developmental biology, and synthetic bioengineering.
Copyright © 2013 Wiley Periodicals, Inc.
Conflict of interest statement
No conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
